
gandum 2









dan gandum lunak (soft wheat). Di daerah yang mempunyai dua musim terdapat
gandum musim panas (hard spring) dan gandum musim dingin (hard winter).
Gandum merah (soft red wheat) dan gandum putih (white wheat)
dikelompokkan sebagai gandum lunak.
Biji gandum memiliki kandungan gizi yang cukup tinggi, diantaranya
karbohidrat 60-80%, protein 10-20%, lemak 2-2,5%, mineral 4-4,5% dan sejumlah
vitamin lainnya (Pomeranz 1971, Šramkováa et al. 2009). Dalam pembuatan
makanan, hal yang harus diperhatikan ialah ketepatan pemakaian jenis tepung
terigu. Tepung terigu dengan kandungan protein 12-14% ideal untuk bahan
baku roti dan mie, 10,5-11,5% untuk biskuit, pastry/pie dan donat. Untuk
gorengan, cake, dan wafer dapat menggunakan gandum dengan kadar protein
8-9%. Jadi, semua tepung terigu belum tentu sesuai dengan produk makanan
yang akan dibuat (Bogasari 2011). Keunggulan mutu protein terigu adalah
kemampuan membentuk gluten yang diperlukan untuk berbagai produk
terutama roti, mie, dan cake. Sifat fisikokimia spesifik ini tidak dimiliki oleh
tepung serealia lainnya. Tulisan ini memaparkan struktur dan komposisi biji
dan nutrisi gandum.
STRUKTUR DAN KOMPOSISI BIJI GANDUM
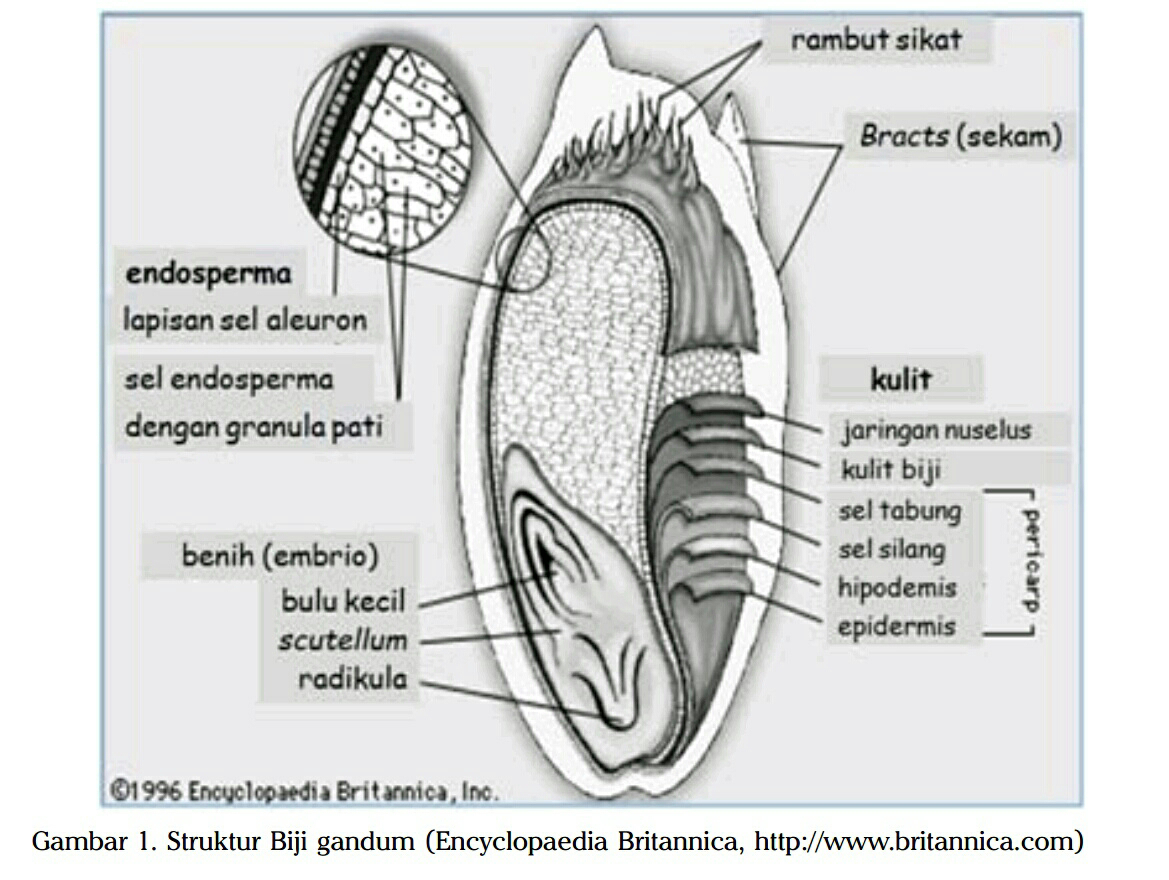
Biji gandum terdiri atas tiga bagian, yaitu lembaga (germ), endosperm, dan
dedak (Eliasson and Larsson 1993). Susunan alami biji gandum adalah dedak
15% (epidemis, epicarp, endocarp, testa dan lapisan aleuron), germ 2,5%, dan
endosperm 82,5% (US Wheat Assosiates 1981). Komposisi ini mendekati
hasil penelitian Belderok et al. (2000) yang melaporkan biji gandum terdiri atas
germ 2-3%, dedak 13-17%, dan endosperm 80-85% basis kering. Komposisi kimia
biji gandum disajikan pada Tabel 1.
Protein biji gandum terkonsentrasi pada bagian germ sekitar 23%, sedangkan
lemak relatif tinggi sekitar 10%, walaupun proporsi dari biji utuh gandum hanya
2-3%. Proporsi endosperm pada biji utuh gandum 80-85%, tidak mengandung
lemak, protein hanya 7%. Pada bagian ini kadar karbohidrat biji gandum sekitar
79%. Bagian dedak dengan proporsi 13-17% mengandung protein 16%, lemak
3%, karbohidrat sekitar 63%. Terlihat dedak biji gandum masih mengandung
gizi tinggi, hal ini berhubungan dengan proses biji gandum menjadi tepung
terigu. Komponen gizi tepung gandum relatif berkurang dari biji utuh, karena
terkikis dan sebagian berada dalam limbah dedak (Tabel 1).
Morfologi biji gandum umumnya terdiri atas kernel berbentuk oval dengan
panjang 6-8 mm dan diameter 2-3 mm, memiliki tekstur yang keras seperti
serealia lainnya. Biji gandum memiliki tiga komponen penting (Gambar 1).
Komponen pertama adalah kulit luar (bran), merupakan kulit luar gandum
dengan proporsi 14,5% dari total keseluruhan biji gandum. Bran terdiri atas lima
lapisan, yaitu epidermis (3,9%), epikarp (0,9%), endokarp (0,9%), testa (0,6%),
dan aleuron (9%). Bran memiliki granulasi yang lebih besar dibanding pollard,
memiliki kandungan protein dan kadar serat tinggi sehingga baik dikonsumsi
ternak besar. Komponen serat kasar pada dedak gandum didominasi oleh
arabino-xylan 66,5% dan selulosa 15% (Lu et al. 2000). Jaroni et al. (1999)
melaporkan konsumsi ransum yang tinggi serat (β-glucan dan arabino-xylan)
akan meningkatkan kekentalan digesta sehingga laju digesta dalam saluran
pencernaan menurun yang berakibat turunnya konsumsi ransum. Oleh sebab
itu, dedak gandum sesuai untuk pakan ternak besar, tetapi untuk ransum ungags
sebaiknya diberi perlakuan pengolahan sebelum digunakan. Perlakuan enzim
kasar asal A. niger dan T. viride pada dedak gandum dapat menurunkan
kandungan serat kasar dan meningkatkan nilai energi metabolis dedak.
pemakaian dedak gandum hasil olahan enzim kasar dalam ransum dapat
memberikan pengaruh terhadap konversi ransum ayam broiler (Ramli et al.
2005).
Epidermis merupakan bagian terluar biji gandum, mengandung banyak
debu yang apabila terkena air akan menjadi liat dan tidak mudah pecah
(Belderok et al. 2000). Fenomena ini yang dimanfaatkan pada penggilingan
gandum menjadi tepung terigu agar lapisan epidermis yang terdapat pada biji
gandum tidak hancur dan tidak mengotori tepung terigu yang dihasilkan.
Kebanyakan protein yang terkandung dalam bran adalah protein larut
(albumin dan globulin).
Komponen kedua adalah endosperma (endosperm), merupakan bagian
terbesar dari biji gandum (80-83%) yang banyak mengandung protein, pati, dan
air. Pada proses penggilingan, bagian ini akan diubah menjadi tepung terigu
dengan tingkat kehalusan tertentu. Pada bagian ini terdapat zat abu yang
kandungannya akan semakin kecil jika mendekati inti dan akan semakin besar
jika mendekati kulit.
Komponen ketiga adalah lembaga (germ), terdapat pada biji gandum
dengan proporsi 2,5-3%. Lembaga merupakan cadangan makanan yang
mengandung banyak lemak dan terdapat bagian yang selnya masih hidup,
bahkan setelah pemanenan. Di sekeliling bagian yang masih hidup terdapat
sedikit molekul glukosa, mineral, protein, dan enzim. Pada kondisi yang lembab
akan terjadi perkecambahan, yaitu biji gandum akan tumbuh menjadi tanaman
yang baru (Kent 1966). Perkecambahan merupakan salah satu hal yang harus
dihindari pada tahap penyimpanan biji gandum. Perkecambahan ini
dipengaruhi oleh beberapa faktor, di antaranya kondisi kelembaban yang tinggi,
suhu yang relatif hangat, dan kandungan oksigen yang tinggi (Singh and Singh
2001).
Intisari gandum adalah lembaga yang merupakan embrio pada tanaman
gandum dengan warna cokelat keemasan dan berbentuk serpihan
(Shellenberger 1971). Murtini et al. (2005) yang menginformasikan dari hasil
penelitiannya, bahwa bila berat biji gandum lebih besar maka kandungan
endosperm tinggi dan tepung yang dihasilkan lebih banyak. Pada produksi
tepung terigu, intisari gandum sering kali dihilangkan pada saat proses
pemurnian biji. Manfaatnya adalah untuk mengantisipasi kandungan minyak
nabati yang tinggi pada intisari gandum, sehingga menghilangkannya akan
mencegah tepung terigu tidak mudah teroksidasi, tidak cepat tengik, dan
memperpanjang umur simpan.
KOMPOSISI GIZI BIJI GANDUM
Komposisi kimia termasuk komponen proksimat gandum relatif tidak berbeda
dengan serealia lainnya. Informasi komposisi kimia proksimat gandum cukup
banyak tersedia. Keragaman data pada masing-masing komponen gizi
dipengaruhi oleh faktor genetik dan lingkungan pertanaman (Pomeranz 1971).
Secara umum, pengaruh suhu tinggi terhadap perkembangan bulir pada serealia
termasuk gandum meliputi laju perkembangan bulir yang lebih cepat,
penurunan bobot bulir, biji keriput, berkurangnya laju akumulasi pati, perubahan
komposisi lipid dan polipeptida (Stone 2001). Komposisi kimia proksimat
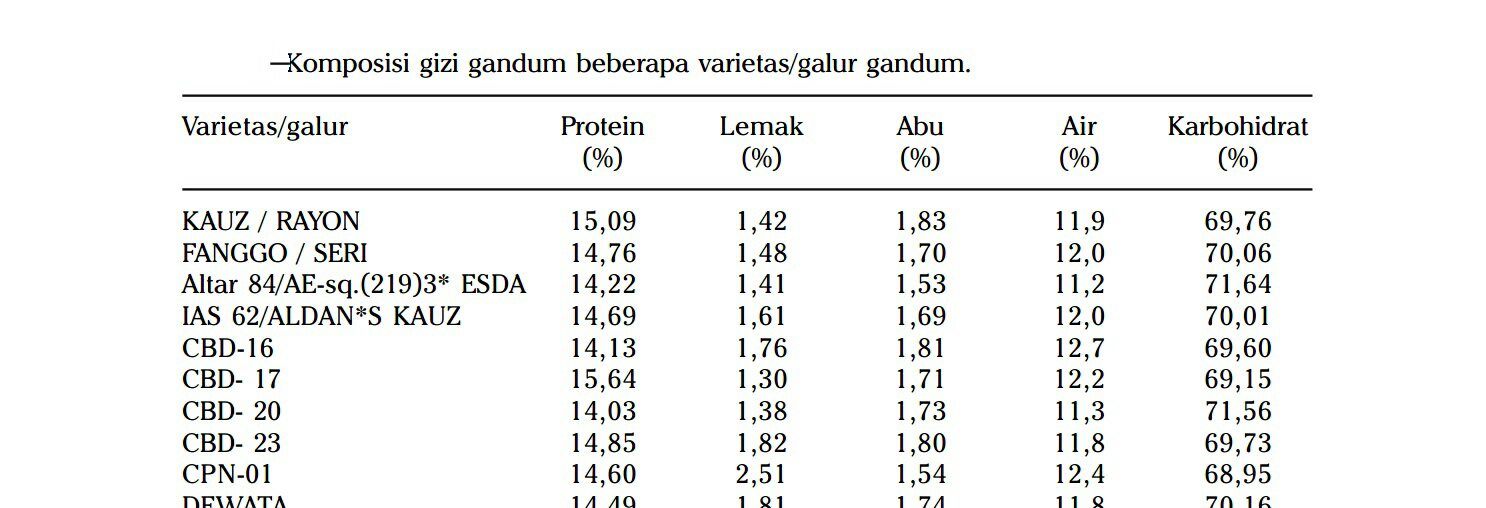
sejumlah galur dan varietas gandum yang telah dilepas Badan Litbang Pertanian
(Nias, Dewata, dan Selayar)
Kadar Abu
Rata-rata kadar abu galur/varietas gandum 1,70% dengan kisaran 1,54-1,83%,
terendah pada CPN-01 dan tertinggi pada Kauz/Rayon. Komponen abu
merupakan sumber mineral pada bahan pangan, tetapi pada kadar tinggi
berpengaruh terhadap tampilan warna tepung terigu yang dihasilkan (Winarno
2002). Kadar abu pada biji gandum akan turun dalam prosesing menjadi tepung
terigu. Misalnya pada varietas Dewata, dari 1,54% pada biji turun menjadi 0,67%
setelah jadi tepung terigu. Pada varietas Nias, 1,36% dari biji menjadi 0,66% setelah
jadi tepung terigu. Pada varietas Selayar, 1,58% dari biji menjadi 0,57% setelah
jadi tepung terigu. Hal ini disebabkan kandungan mineral/abu terkonsentrasi
60-65% pada aleuron layer, sisanya pada endosperm. Pada saat prosesing, bagian
aleuron terkikis terikut menjadi limbah. Sejumlah galur/varietas gandum
menunjukkan beragam kadar protein, abu, lemak dan berat gluten (Tabel 3).
Protein
Kadar protein galur/varietas gandum yang diteliti berkisar antara 12,7-16,8%,
terendah pada galur PICUS/4/CS5A dan tertinggi pada galur BAW 898 (Suarni
dan Hamdani 2009). Kadar abu berkisar antara 1,4-2,6, terendah pada varietas
Dewata dan tertinggi pada galur PASTOR/2*SITTA. Tingginya kadar abu galur/
varietas gandum ini menunjukkan tingginya kandungan mineralnya.
Kandungan lemak galur/varietas gandum berkisar antara 1,44-2,05%, terendah
pada galur KAUZ/WEAVER dan tertinggi pada galur VEE/PJN//2*TUI. Berat gluten
gandum berkisar antara 25,2-41,4%, terendah pada galur W462/VEE/KOEL/3/
PEG/MRL/BUC dan tertinggi pada galur PFAU/WEAVER.
Kadar protein biji gandum dengan varietas yang sama menunjukkan
konsentrasi yang berbeda, hal ini dapat diakibatkan perbedaan iklim, kondisi
lahan, pertumbuhan yang kurang optimal. Sehubungan dengan hal ini ,
kadar protein varietas Selayar dan Dewata lebih rendah dibanding hasil penelitian
di atas (Sihotang et al. 2015). Sebelumnya Murtini et al. (2005) meneliti protein
biji gandum varietas Selayar, Nias dan Dewata tidak beda jauh dengan hasil
penelitian Suarni dan Hamdani (2009).
Protein merupakan salah satu cadangan makanan yang terdapat dalam biji
gandum dan banyak terdapat di bagian endosperm biji. Protein berguna
menunjang pertumbuhan biji selama proses berkecambah, sehingga jenis
protein yang dibutuhkan selama proses perkecambahan adalah protein terlarut
(dapat dihidrolisis dengan mudah oleh protease). Biji gandum mengandung
beberapa jenis protein penting, yaitu albumin, globulin, gliadin, dan prolamin,
yang tersimpan pada endosperm sebagai cadangan makanan dan sewaktuwaktu dirombak untuk proses perkecambahan (Sramkovaa et al. 2009).
Kandungan protein merupakan salah satu tolok ukur yang sangat penting
untuk mengetahui katagori gandum. Pada biji gandum terkandung protein 10-
20%. Rata-rata kadar protein biji gandum yang diteliti adalah 14,65%, dengan
kisaran 14,03-15,64%, tertinggi pada CBD-17 dan terendah pada CBD-20 (Tabel
2). Angka ini relatif sama dengan kadar protein gandum varietas Selayar, Nias,
dan Dewata berkisar antara 12,9-14,5%
Perubahan kadar protein dari biji gandum menjadi tepung terigu relatif kecil.
Misalnya varietas Dewata, dari 14,0% pada biji menjadi 13,7% pada tepung terigu.
Pada varietas Selayar dari 13,6% pada biji menjadi 13,1% pada tepung terigu.
Pada varietas Nias dari 12,9% pada biji menjadi 13,4% pada tepung terigu (Tabel
3). Perubahan komposisi nutrisi biji gandum setelah menjadi tepung terigu
akibat proses penepungan, sehingga kehilangan kandungan serat pangan,
fitosterol pada produk tepung terigu (Pomeranz 1971). Perubahan komposisi
protein dan nutrisi lainnya terjadi sejak tahap pengisian biji hingga panen, dan
umur panen berpengaruh terhadap kadar protein biji (Daniel and Triboi 2002).
Salah satu faktor yang menentukan mutu protein bahan pangan adalah
komponen asam amino penyusunnya. Kandungan asam amino esensial relatif
lebih tinggi pada terigu dibanding tepung sorgum (Tabel 4). Pada terigu, asam
amino lisin 0,38%, lebih tinggi dibanding tepung sorgum yang hanya 0,18%.
Kadar asam amino fenilalanin tepung terigu adalah 0,61%, lebih tinggi dibanding
tepung sorgum dengan kadar 0,18%. Kadar prolin pada terigu adalah 1,51%,
lebih tinggi dari tepung sorgum yang hanya 0,29%.
Kadar asam glutamat terigu adalah 3,83%, lebih tinggi dibanding tepung
sorgum yang hanya 1,58%. Meskipun asam glutamat bukan termasuk asam
amino esensial, namun berpengaruh terhadap sifat sensori produk olahan,
terutama dari segi rasa. Hal ini ditunjukkan oleh hasil uji organoleptik dari segi
rasa roti tawar dengan bahan tepung sorgum yang hanya dapat mensubstitusi
terigu hingga 20% (Suarni dan Patong 2002).
Dalam pembuatan makanan, hal yang perlu diperhatikan adalah ketepatan
pemakaian jenis tepung terigu. Tepung terigu dengan kadar protein 12-14% ideal
untuk bahan roti dan mie, 10,5-11,5% untuk biskuit, pastry dan donat, sedangkan
untuk gorengan, cake dan wafer sebaiknya dengan kadar protein 8-9%.
Lemak
Lemak dalam bahan makanan merupakan komponen esensial yang dibutuhkan
tubuh, tetapi berpengaruh terhadap umur simpan bahan pangan ini . Biji
gandum mengandung lemak 2-2,5% (bk) dengan konsentrasi berkisar antara
25-30% pada bagian germ (Winarno 2002, Didin 2008). Kisaran kadar lemak
1,82-2,51% dengan rata-rata 1,65 (Tabel 2). Lemak biji gandum tediri atas
campuran trigliserida, yaitu senyawa gliserol dan tiga asam lemak berupa
fosfatidilkolin kolin, etanolamin fosfatidilkolin, dan fosfatidilkolin serin, serta 32
turunan lyosophosphatidyl, di mana ada satu grup hidroksil bebas pada bagian
gliserol. Komposisi lemak gandum sangat berpengaruh pada kualitas tepung.
Biji gandum yang berkualitas tinggi antara lain memiliki kandungan lemak yang
rendah. Biji gandum yang dikecambahkan selama beberapa jam akan
menghasilkan tepung rendah lemak dan dapat digunakan sebagai makanan
diet. Dalam keadaan tertentu, biji gandum perlu dikecambahkan untuk
menghasilkan tepung rendah lemak yang diolah menjadi makanan diet. Selain
itu, selama proses ini terjadi penurunan kandungan senyawa antinutrisi,
seperti tripsin inhibitor, tanin, pentosan, dan asam fitat (Handoyo et al. 2006,
Handoyo 2008). Perkecambahan selama beberapa jam memicu
menurunnya kandungan lemak pada biji gandum DNS 14 (jenis Dark Northen
Spring). Kandungan lemak tertinggi terdapat pada tanpa perkecambahan
(kontrol) sebesar 2,11% dan terendah pada lama perkecambahan 32 jam yaitu
1,84% (Indaryati 2011). Selama perkecambahan, lemak terhidrolisis menjadi
asam lemak dan gliserol dengan bantuan enzim lipase. Menurut Pranoto et al.
(1990), hasil perombakan lemak berupa asam lemak bebas dan gliserol
kemudian dipindahkan ke embrio. Menurut Miyake et al. (2004),
perkecambahan dapat menurunkan kadar lemak pada buck wheat dan
gandum. Semakin lama waktu perkecambahan, semakin berkurang
ketersediaan lemak dalam biji gandum.
Bagian lembaga biji gandum mengandung protein dan lemak bermutu tinggi
sebagai pangan fungsional (Matz 1992). Lemak lembaga gandum terdapat pada
komponen fitosterol 1,3-1,7% (Formo et al. 1979). Marliyati et al. (2005) telah
mengekstrak lemak lembaga gandum dengan nisbah pelarut heksan : etanol
1:3 (v/v) dengan rendemen 15,8%. Mengekstrak komponen fitosterol dari lemak
ini menghasilkan rendemen tertinggi dengan nisbah pelarut heksan :
etanol 82:18 (v/v) yaitu 1,37% atau 11,70% terhadap lipida. Suplementasi sterol
lembaga gandum ke dalam margarin dapat mengimbangi kolesterol yang
terkandung dalam bahan makanan ini (Marliyati et al. 2010).
Fitosterol mempunyai sifat antiaterogenik, sehingga mengonsumsi fitosterol
dalam jumlah yang banyak dapat menekan penyerapan kolesterol dalam tubuh
sehingga akan meningkatkan ekskresinya (Hui 1996). lntervensi secara klinis
rnenunjukkan kadar kolesterol total dan LDL (Low Density Lipid) dapat
mencegah penyakit jantung coroner (Cleghorn et al. 2003).
Karbohidrat
Gandum merupakan komoditas serealia sumber karbohidrat. Sebagian besar
komposisi karbohidrat gandum adalah pati. Gandum dan serealia lainnya
menyimpan energi dalam bentuk pati. Jumlah pati yang terkandung dalam
sebutir gandum bervariasi antara 60-75% dari total bobot kering. Kekurangan
pati gandum adalah tidak dapat diekstrak seperti pati jagung dan sorgum, karena
matriks proteinnya sangat kuat, bahkan membentuk gluten yang apabila
ditambahkan air.
Pati gandum terbentuk pada benih dalam bentuk butiran, memiliki dua
jenis granula pati, besar (25-40 μm) lenticular dan kecil (5-10 μm) yang bulat.
Menurut Stoddard (1999), pati dengan ukuran granula yang kecil baik untuk
dijadikan bahan baku makanan. Pati gandum dengan ukuran granula yang
sempit atau seragam akan menghasilkan produk yang lebih baik. Granula
lenticular terbentuk selama 15 hari pertama setelah penyerbukan. Butiran kecil,
representating sekitar 88% dari total butiran, muncul 10-30 hari setelah
penyerbukan (Belderok et al. 2000). Glenn dan Saunders (1990) mengamati
bentuk dan ukuran granula pati gandum menggunakan Scanning Electron
Microscopy (SEM) beberapa genotipe gandum. Hasilnya terdapat perbedaan
ukuran, tetapi hanya memiliki dua jenis granula pati.
Ukuran granula pati gandum, jagung, dan sorgum dengan menggunakan
SEM dapat dilihat pada Gambar 2. Terlihat perbedaan bentuk dan ukuran antara
granula pati terigu, pati jagung dan pati sorgum. Bentuk granula pati varietas
gandum Selayar, Nias, dan Dewata relatif sama, hanya ukurannya yang berbeda
(Suarni et al. 2009).
Secara kimiawi, pati merupakan polimer glukosa yaitu amilosa dan
amilopektin. Pati gandum yang normal biasanya mengandung 20-30% amilosa
dan 70-80% amilopektin (Belderok et al. 2000, Suarni dan Hamdani 2009). Pati
biji gandum terbentuk dari dua jenis polimer glukosa yaitu amilosa dan
amilopektin (Satorre and Slafer 1999). Amilosa merupakan rantai unit-unit Dglukosa yang panjang dan tidak bercabang, digabungkan oleh ikatan α(1→4),
sedangkan amilopektin memiliki struktur bercabang. Ikatan glikosidik yang
menggabungkan residu glukosa yang berdekatan dalam rantai amilopektin ialah
ikatan α(1→4), tetapi titik percabangan amilopektin merupakan ikatan α(1→6).
Bobot molekul amilosa dan amilopektin bergantung pada asal sumber
botaninya. Amilosa merupakan komponen dengan rantai lurus, sedangkan
amilopektin adalah komponen dengan rantai bercabang. Amilosa merupakan
polisakarida berantai lurus berbentuk heliks dengan ikatan glikosidik α-1,4
(Gambar 3). Jumlah molekul glukosa pada rantai amilosa bervariasi antara 250-
350 unit. Amilopektin merupakan polisakarida bercabang dengan ikatan
glikosidik a-1,4 pada rantai lurus dan ikatan α-1,6 pada percabangan (Gambar
4). Titik percabangan amilopektin lebih banyak dibanding amilosa (Dziedzic
and Kearsley 1995).
Salah satu komponen karbohidrat gandum yang bersifat pangan fungsional
adalah serat pangan yang sampai saat ini paling banyak digunakan dalam
makanan fungsional. Serat dedak gandum dan gum adalah contoh serat
makanan yang sering ditambahkan ke dalam makanan fungsional. Serat
makanan yang larut dalam air seperti polydextrose digunakan dalam minuman
fungsional. Manfaat fisiologis produk yang diberi serat makanan antara lain
mengatur fungsi-fungsi usus, mencegah penyakit divertikulosis, konstipasi,
mengendalikan kolesterol darah, mengatur kadar gula darah, mencegah
obesitas dan mengurangi risiko kanker kolon (Irawan dan Wijaya 2002).
Keberadaan serat makanan dalam menu sehari-hari dapat menjaga dan
meningkatkan fungsi saluran cerna serta menjaga kesehatan tubuh, terutama
untuk menghindari berbagai penyakit degenaratif, seperti obesitas, diabetes
melitus, dan penyakit kardiovaskuler
Berdasarkan kemampuannya untuk larut dalam air, serat makanan
dikelompokkan ke dalam serat larut (soluble fiber) dan serat tidak larut (insoluble
fiber). Serat larut meliputi pektin, gum, α-glukan, selulosa, hemiselulosa, dan
lignin (Wildman 2000). Diperkirakan sepertiga serat makanan total (total dietary
fiber/TDF) adalah serat makanan larut, sedangkan yang terbanyak adalah serat
tidak larut (Gordon 1989). Nilai kecukupan asupan serat makanan yang
dianjurkan untuk orang negara kita dewasa adalah 20-35 g/hari. Walaupun nilai
kecukupan yang dianjurkan cukup tinggi, hasil survei menunjukkan asupan
rata-rata serat makanan orang dewasa negara kita hanya 10,5 g/hari (Direktorat
Gizi warga 2000).
Mineral dan Vitamin
Bahan makanan yang dikonsumsi termasuk terigu kekurangan zat besi, Zn, dan
pro. vit A (Sramkovaa et al. 2009). Rodriguez et al. (2011) menginformasikan
bahwa biji gandum memiliki kandungan mineral fosfor (2370 ± 333 mg/kg),
natrium (102 ± 52 mg/kg), kalium (4363 ± 386mg/kg), kalsium (351 ±62 mg/
kg), magnesium (1163 ±155 mg/kg), besi (40,0 ±5,5 mg/kg), tembaga (2,68 ±
0,93 mg/kg), seng (32,1 ±2,9 mg/kg), mangan (22,1 ±3,5 mg/kg), dan selenium
(67,7 ± 40,4 ìg/kg). Gandum kaya vitamin B kompleks. Di antara sumber vitamin
B, kadar riboflavin lebih tinggi dibandingkan beras pecah kulit, tiamin setara dengan
beras pecah kulit, sedangkan kadar niasin tertinggi pada gandum, dan riboflavin
setara dengan sorgum tetapi lebih rendah dibanding jagung (Tabel 5). Komoditas
serealia termasuk sumber mineral kalsium relatif tinggi dengan kisaran 25-35
mg/100g, gandum setara dengan beras pecah kulit, sorgum terendah dan
jewawut tertinggi.
Kelebihan sorgum adalah mengandung kadar besi yang lebih tinggi
dibanding serealia lainnya. Kandungan besi gandum adalah 3,5 mg/100 g masih
termasuk tinggi, tetapi akan turun drastis dalam bentuk terigu, akibat terkikis
pada setiap tahapan proses pengolahan dari gandum menjadi terigu. Hal ini
juga terjadi pada komponen mineral dan vitamin lainnya.
SIFAT FISIKOKIMIA GANDUM DAN TERIGU
Sifat fisiko kimia gandum dan terigu perlu diketahui karena merupakan landasan
dalam menentukan produk yang akan dihasilkan. Sifat fisiko kimia gandum
dan terigu dari varietas Selayar, Dewata dan Nias disajikan pada Tabel 6.
Terigu adalah produk dari biji gandum setelah melalui proses cukup panjang.
Pada pabrik pengolahan gandum menjadi terigu, kegiatan uji sifat fisiko kimia
sampel setiap produksi merupakan keharusan sehingga menjadi pekerjaan
rutin. Kualitas tepung terigu dipengaruhi oleh beberapa parameter seperti
moisture (kadar air), ash (kadar abu), dan beberapa parameter fisik lainnya
seperti water absorption, development time, stability, dan parameter fisikokimia
lainnya (Glenn and Saunders 1990, Uthayakumaran and Lukow 2003)
Moisture adalah kadar air tepung terigu yang mempengaruhi kualitas tepung.
Bila jumlah kadar air melebihi standar maksimum maka memicu terjadinya
penurunan daya simpan tepung terigu karena cepat rusak, berjamur, dan bau
apek.
Ash adalah kadar abu pada tepung terigu yang mempengaruhi proses dan
hasil akhir produk, antara lain warna produk (warna crumb pada roti, warna
mie) dan tingkat kestabilan adonan. Semakin tinggi kadar abu semakin buruk
kualitas tepung, sebaliknya semakin rendah kadar abu semakin baik kualitas
tepung. Tingginya kadar abu suatu bahan menunjukkan tingginya komponen
mineral.
Water Absorption (daya serap air) adalah kemampuan tepung terigu
menyerap air. Kemampuan daya serap air tepung terigu berkurang bila kadar
air dalam tepung (moisture) terlalu tinggi atau disimpan pada tempat yang
lembab. Daya serap air bergantung pada produk yang akan dihasilkan, dalam
pembuatan roti umumnya diperlukan daya serap air yang lebih tinggi dibandingkan
pembuatan mie dan biskuit.
Developing Time adalah kecepatan tepung terigu dalam pencapaian
keadaan develop (kalis). Bila waktu pengadukan adonan kurang sempurna,
disebut under mixing, memicu volume adonan tidak maksimal sehingga
tekstur/remah roti kasar, roti terlalu kenyal, aroma roti asam, roti cepat keras,
permukaan kulit roti pecah dan tebal. Sebaliknya, bila kelebihan pengadukan
adonan, disebut over mixing, memicu volume roti melebar, kurang
mengembang, remah roti kasar, warna kulit roti pucat, permukaan roti mengecil,
permukaan kulit roti terdapat banyak gelembung dan roti kurang kenyal.
Stability adalah kemampuan tepung terigu untuk menahan stabilitas adonan
agar tetap sempurna meskipun telah melewati waktu develop (kalis). Stabilitas
tepung pada adonan dipengaruhi oleh beberapa hal, antara lain kandungan
protein, kualitas protein, dan zat tambahan (additive).
Gluten
Gluten merupakan campuran protein antara dua jenis protein gandum yaitu
glutenin dan gliadin. Protein dibedakan menjadi (1) larut yaitu albumin-globulin
dan (2) tidak larut yaitu gliadin-glutenin, perbandingan senyawaan ini
dalam kondisi yang baik untuk membentuk gluten. Glutenin memberikan sifat
yang tegar dan gliadin memberikan sifat yang lengket, sehingga mampu
memerangkap gas yang terbentuk selama proses pengembangan adonan
(Winarno 2002). Dias et al. (2011) menjelaskan bahwa gluten dibentuk dari
gliadin dan glutenin, gluten mempunyai peranan penting dalam pembentukan
struktur, secara fungsional dapat meningkatkan nilai baking expansion karena
bersifat hidrofilik. Gluten dapat merenggangkan ikatan antar molekul sehingga
air akan masuk ke dalam molekul pati, akibatnya terjadi peningkatan volume
dan pengembangan granula pati pada saat pemanggangan, serta kemampuan
gluten mengikat molekul air dengan ikatan hidrogen yang kuat, dapat
meningkatkan daya kembang produk akhir.
Kualitas produk bakery/roti, mi dan sejenisnya bergantung pada kualitas
terigu. Terigu mempunyai kelebihan dibanding tepung yang lainnya, terletak
pada sifat pembentukan gluten. Gluten bersama pati gandum akan membentuk
struktur dinding sel (building block) yang menghasilkan remah produk. Sifat
spesifik ini kurang dimiliki oleh serealia lainnya, termasuk jagung, sorgum,
jewawut dan padi, tetapi menjadikannya tidak sesuai dengan penderita alergi
gluten.
Gluten memicu penyakit intoleransi terhadap gluten (celiac disease).
Kondisi ini ditandai oleh terjadinya radang mukosa usus halus sehingga
tidak dapat berfungsi secara normal. Gluten terdapat pada gandum, gandum
hitam, dan barley. Untuk menghindari konsumsi gluten dapat mengkonsumsi
produk lain yang berasal dari beras, jagung, dan sorgum (Winarno 2002, Steven
2004). Pemberian enzim protease memicu penurunan kandungan
protein alergenik pada tepung terigu. Perombakan oleh enzim protease juga
terhadap protein alergenik menjadi asam amino bebas dan protein sederhana,
maka kandungannya lebih rendah. Tepung gandum rendah protein alergenik
hasil dari metode pre-germinated wheat flour sesuai bagi penderita alergi,
sehingga dapat mengonsumsi makanan yang dibuat dari gandum (Handoyo
2008).
Penelitian Suarni dan Zakir (2001) menunjukkan kandungan gluten sorgum
sangat rendah <1% dengan mutu yang kurang baik, sehingga tepung sorgum
hanya mampu mensubstitusi 15-20% terigu untuk produk roti dan sejenisnya.
Kandungan gluten pada gandum dan terigu dari beberapa galur dan varietas
dapat dilihat pada (Tabel 3 dan 6).
Kemampuan daya bentuk produk dari terigu ditentukan oleh mutu dan
jumlah glutennya. Jenis terigu yang dibuat dari gandum keras (hard wheat)
mengandung protein yang bermutu baik (>10,5%) sesuai untuk pembuatan
roti. Jenis terigu dari gandum lunak (soft wheat) dengan kandungan protein
<10% digunakan untuk membuat cake, cookies, pastel (U.S. Wheat Associates
1983). Gluten gandum dapat diekstrak dan telah dikomersialkan, dibutuhkan
pada adonan berbasis tepung yang kurang memiliki gluten, dengan produk
yang memerlukan elastisitas adonan dan pengembangan volume olahan pada
pemanggangan. Gluten gandum (wheat gluten/WG) diekstrak dari bagian
endosperm dengan komposisi protein-lemak-pati. Gluten gandum komersial
memiliki komposisi 72,5% protein (77,5% berat kering), 5,7% lemak, 0,7% abu,
6,4% air, karbohidrat, pati, dan komponen lainnya (Clodualdo et al. 1994).
PENUTUP
Informasi karakter struktur biji, komposisi kimia, sifat fisiko kimia dan fungsional
serta mutu nutrisi gandum setiap varietas diperlukan sebagai rujukan oleh
pemulia gandum dalam merakit varietas unggul. Hal ini juga diperlukan untuk
diversifikasi pangan dan sebagai landasan bagi industri pangan dalam memilih
bahan baku/varietas dan pengolahan yang sesuai dengan produk yang
diinginkan. Dedak gandum mengandung serat kasar yang tinggi, berpotensi
sebagai bahan pakan ternak besar, sedangkan untuk ternak unggas
memerlukan perlakuan pengolahan sebelum digunakan.
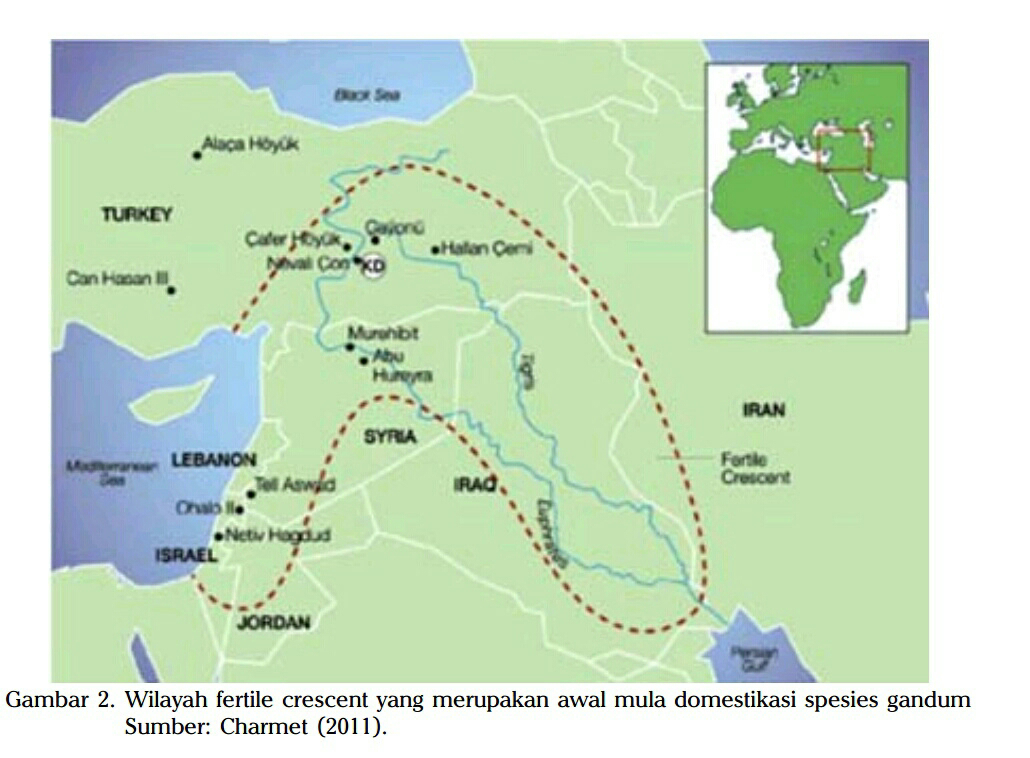
Gandum di negara kita merupakan tanaman pendatang, karena komoditas ini
merupakan serealia dari daerah yang memiliki suhu musim panas yang hangat
dan musim dingin yang dingin, seperti wilayah beriklim dingin (temperate),
mediterania, subtropis, dan wilayah-wilayah tropis dengan ketinggian di atas
1.000 m dpl (Percival 1921, Elias 1995, OECD 1999, Nevo et al. 2002). Gandum di
daerah subtropis berdasarkan waktu tanamnya ada dua jenis, yaitu gandum
musim dingin (winter wheat) dan gandum musim panas (spring wheat).
Gandum yang ditanam di daerah semi-arid di kawasan mediteran yang bersuhu
dingin adalah jenis spring wheat. Seiring dengan perkembangan program
pemuliaan tanaman gandum, saat ini gandum sudah dapat dikembangkan di
negara kita yang beriklim tropis (Elias 1995, OECD 1999, Hamdani et al. 2002,
Shewry 2009).
Pada awal evolusinya, gandum merupakan tanaman diploid yang kemudian
berkembang menjadi tanaman poliploid melalui proses persilangan alami
dengan kerabat liarnya sejak ribuan tahun yang lalu. Gandum yang saat ini
dikembangkan di seluruh dunia 95% adalah gandum hexaploid (Triticum
aestivum L., 2n=6x=42, AABBDD), atau dikenal sebagai gandum roti (bread
wheat), dan 5% sisanya adalah gandum tetraploid (Triticum turgidum L.,
2n=4x=28, AABB) yang lebih dikenal dengan gandum durum (durum wheat)
(Nevo et al. 2002, Shewry 2009). Gandum durum merupakan bahan baku pasta,
seperti spageti dan macaroni, sedangkan gandum biasa merupakan bahan
baku roti, cake, dan camilan.
Tulisan ini membahas morfologi dan tahap pertumbuhan gandum yang
diharapkan dapat menambah khasanah ilmu pengetahuan tanaman pangan ini.
MORFOLOGI
Gandum durum dan gandum roti, secara genetik berbeda namun secara
morfologi mirip, yang membedakan adalah kandungan proteinnya. Gandum
durum memiliki kandungan protein yang lebih tinggi dibandingkan gandum roti (Pauly
et al. 2013). Gandum merupakan tanaman kelompok serealia, satu famili
dengan padi, jagung, hanjeli, dan sorgum. Dalam sistem taksonomi tumbuhan,
gandum termasuk dalam keluarga Poaceae atau lebih dikenal sebagai Gramineae
(rumput-rumputan), dengan ciri khas berakar serabut, batang berbuku, dan daun
sejajar dengan tulang daun. Secara umum morfologi tanaman gandum terdiri
atas akar, batang, daun, anakan, bunga dan biji
Akar
Sebagaimana tanaman serealia lainnya, sistem perakaran gandum adalah
perakaran serabut. Terdapat dua tipe perakaran gandum, yaitu akar primer
(seminal root) dan akar skunder (nodal root). Akar primer terdiri atas radikula
(radicle) dan akar seminal lateral (lateral seminal roots) (Gambar 2). Akar primer
berkembang dari primordial akar yang tumbuh pada saat biji gandum
berkecambah, pada saat masih embrio, setidaknya terdapat enam akar
primordial, yang terdiri atas satu akar radikula (radical) dan dua pasang akar
seminal lateral. Pada saat perkecambahan akar primordial (radikula dan
seminal) menembus koleoriza dan akan tumbuh memanjang hingga kurang
lebih 2 m, akar kemudian digantikan oleh akar skunder (crown/adventitious)
(Gambar 3) (Percival 1921, Kirby 2002).
Akar sekunder berkembang pada saat tanaman mulai membentuk anakan
atau telah memiliki empat daun. Akar skunder tumbuh pada buku mahkota
(crown nodes) yang terletak pada buku ke-3-7 paling bawah dari tunas utama
dan anakan. Pada buku yang lebih atas, setiap buku terdapat 5-6 akar (Percival
1921, Kirby 2002). Akar akan memanjang hingga kedalaman 2 m, bergantung
pada jenis tanah (Kirby 2002, Australian Government 2008). Pada musim dingin
(winter wheat), perakaran gandum bisa mencapai 2,2 m, namun pada musim
semi (spring wheat) hanya sekitar 1,1 m (Kirby 2002, Thorup-Kristensen et al.
2009). Hal ini kemungkinan disebabkan oleh umur gandum musim dingin lebih
lambat, yaitu 280-350 hari, sedangkan gandum musim panas 120-145 hari
(Weaver 1926, FAO 2013). Kepadatan perakaran gandum dalam tanah kurang
dari 0,5 cm3
pada setiap 1cm3
tanah (Atta et al. 2013). Jaringan meristem pada
akar gandum terletak pada 2-10 mm di setiap ujung akar (Anderson and Garlinge
2000). Jumlah akar skunder akan terus bertambah seiring dengan pertambahan
jumlah anakan (Weaver 1926). Laju pertumbuhan akar 1,0-1,5 cm/hari (Anderson
and Garlinge 2000). Anakan akan membentuk akar sendiri pada saat memiliki
setidaknya tiga daun (Australian Government 2008).
Batang
Batang gandum berupa jerami yang tegak, berbentuk silinder dan memiliki
permukaan yang halus, tersusun atas beberapa buku dan ruas (Gambar 4).
Ruas dan buku pada tanaman gandum berkisar antara 8-16. Buku batang
gandum umumnya keras, berfungsi sebagai tempat tumbuhnya daun, akar,
anakan dan malai, juga sebagai perantara keluar masuknya hara tanaman dan
fotosintat. Jarak antara dua buku disebut ruas. Sebagaimana ruas-ruas jerami
pada batang padi, ruas pada batang gandum juga berlubang di tengahnya,
namun pada beberapa varietas ada yang berisi empulur yang lembut.
Batang gandum terbungkus oleh pelepah daun guna menunjang batang
agar tetap tegak sehingga tidak mudah rebah. Ruas pada batang bawah
umumnya lebih pendek dari ruas yang ada di ujung tanaman, bahkan
berhimpitan satu sama lain, membentuk mahkota (crown). Mahkota ini tersusun
atas 8-14 buku dan ruas, ruas pada mahkota hanya berukuran beberapa
millimeter, sedangkan 4-7 ruas terakhir akan tumbuh lebih panjang. Ruas paling
panjang pada batang gandum adalah ruas terakhir (peduncle), yang berfungsi
menopang malai. Pemanjangan ruas terakhir berhenti pada saat anthesis. Ukuran
batang bergantung pada varietas dan lingkungan tumbuh. Pemanjangan ruas
dimulai pada saat tanaman mulai dewasa. Batang juga berfungsi sebagai tempat
penyimpanan cadangan fotosintat untuk pengisian biji. Pada saat malai mulai
muncul, kandungan karbohidrat pada batang gandum mencapai 25-40% dari
bahan kering total (Percival 1921, Anderson and Garlinge 2000, Kirby 2002,
Australian Government 2008).
Daun
Daun gandum berbentuk pita sejajar tulang daun, tersusun atas helai daun
(leaf blade), pelepah daun (leaf seath), ligula (ligule) dan aurikel (auricle)
(Gambar. 5). Bagian dasar pelepah daun melekat pada buku dan menyelimuti
batang. Pelepah daun berfungsi melindungi batang dari cuaca ekstrim dan
menopang batang agar tidak mudah rebah. Batang gandum bagian bawah
tertutup oleh pelepah yang saling tumpang tindih, sehingga batang tidak terlihat.
Namun pada ruas terakhir, pelepah daun akan menutupi bakal malai sebelum
malai pecah. Setelah malai pecah/muncul dan ruas terakhir memanjang, hanya
sebagian batang yang akan tertutup oleh pelepah (Percival 1921, Anderson and
Garlinge 2000, Kirby 2002).
Pada ujung pelepah daun terdapat helai daun. Helai daun gandum memiliki
permukaan yang licin, kadang terdapat sedikit bulu tipis. Pada beberapa varietas,
tepi daun juga berambut. Bagian bawah daun umumnya lebih halus dari sisi
bagian atas. Ukuran daun beragam, semakin ke atas semakin lebar, namun
akan memendek pada lima daun terakhir. Rata-rata daun gandum berukuran
5-10 cm. Daun yang terakhir muncul adalah daun bendera. Pada pertemuan
antara daun dan pelepah terdapat aurikel. Aurikel berbentuk kurva yang melekat
pada dasar setiap helai daun, berwarna hijau muda atau merah muda, dengan
bagian tepi berumbai atau juga berambut, aurikel melekat pelepah dengan
batang. Pada daun yang lebih atas, aurikel melekat lebih kuat dari padadaun
bagian bawah. Ligula pada daun gandum merupakan struktur membrane tipis
yang melekat pada batang, berfungsi mencegah air hujan, debu dan serangga
masuk ke dalam pelepah dan batang gandum. Ligula umumnya tidak berwarna
dengan pinggir yang tidak rata dan sedikit berjumbai. Ukuran ligula berkisar
antara 3-4 mm, semakin ke bawah letak daun semakin pendek ukuran ligula.
(Percival 1921, Anderson and Garlinge 2000, Kirby 2002, Australian Government
2008).
Anakan
Anakan tanaman gandum merupakan cabang lateral yang tumbuh dari pangkal
daun pada batang utama. Anakan memiliki struktur yang sama dengan batang
utama dan membentuk malai, meski tidak semua anakan mampu menghasilkan
malai. Anakan yang terbentuk umumnya sejumlah pangkal daun yang terdapat
pada buku mahkota. Anakan yang terbentuk dari batang utama disebut anakan
primer (primary tiller), yang menghasilkan anakan skunder (scondary tiller).
Pada bagian koleoptil juga kadang tumbuh anakan, yang disebut anakan
koleoptil (colleoptile tiller) (Gambar 6) (Anderson and Garlinge 2000, Kirby 2002,
Australian Government 2008).
Anakan umumnya mulai tumbuh pada saat daun ke-3 telah berkembang
sempurna dan daun ke-4 muncul dari batang utama. Anakan pertama (T1)
tumbuh di bawah ligula daun pertama, begitu seterusnya. Anakan primer
mampu membentuk anakan sendiri yang disebut dengan anakan skunder.
Pertumbuhan anakan ini terhenti setelah terjadi perpanjangan ruas. Jumlah
anakan bergantung pada varietas dan kondisi lingkungan. Jumlah anakan
produktif gandum musim dingin pada kondisi optimal adalah 2-3 batang per
tanaman. Pada kondisi suhu hangat, jumlah anakan yang muncul akan lebih
banyak dibandingkan suhu dingin. Di negara kita dengan suhu yang lebih hangat,
jumlah anakan produktifnya rata-rata 4 batang. Anakan juga memberikan
kontribusi bagi batang utama dalam menghasilkan karbohidrat dan hasil biji.
Bahkan pada kondisi tercekam, anakan sangat berperan dalam menguragi
kerugian hasil, terutama bila batang utama gagal membentuk malai atau rusak.
Jumlah populasi optimal setiap 930 cm2
adalah 30-35 tanaman dengan 70
anakan. Namun pada kondisi kekurangan nutrisi, anakan cenderung tidak
berhasil membentuk malai karena terjadi kompetisi antara batang utama dan
anakan dalam mendapatkan cahaya dan unsur hara serta fotosintat (Percival
1921, Anderson and Garlinge 2000, Kirby 2002, Conley et al. 2003, Australian
Government 2008, Puspita et al. 2013, Wahyu et al. 2013, Wirawan et al. 2013).
Bunga
Pembungaan pada tanaman gandum bersifat determinate, artinya pertumbuhan
vegetatif terhenti pada saat pembungaan. Bunga gandum merupakan
sekelompok bunga yang tersusun dalam malai (ear/spike) (Gambar 6). Pada
setiap malai terdapat beberapa spikelet (spikelet) (Gambar 7), dan setiap spikelet
terdiri atas beberapa bunga tunggal (floret) (Gambar 8) (Percival 1921, de Vreis
1971, Allan 1980, Australian Government 2008, Kirby 2002, Anderson and Garlinge
2000).
Malai gandum tersusun atas spikelet dan tangkai malai (rachis). Pada tangkai
malai utama terdapat beberapa ruas yang pendek sebagai tempat tumbuhnya
spikelet. Terdapat dua baris spikelet pada tangkai malai utama (main axis/rachis),
yang tersusun saling berhadapan. Distribusi spikelet beragam dari sangat rapat
hingga longgar, bergantung pada varietas. Beberapa varietas memiliki spikelet
yang longgar di bagian pangkal dan sangat rapat pada ujungnya. Spikelet yang
berada paling ujung malai disebut spikelet terminal (terminal spikelet). Spikelet
terminal pada beberapa varietas tidak berkembang dengan baik, bahkan tidak
ada, selain itu juga ditemukan spikelet yang tidak berkembang normal pada
pangkal malai. Dalam setiap malai terdapat 5-30 spikelet (Percival 1921, Allan
1980, Australian Government 2008, Kirby 2002, Anderson and Garlinge 2000).
Spikelet merupakan kumpulan dari bunga tunggal (floret), yang tersusun
pada tangkai malai skunder (sub-rachis/rachilla). Setiap spikelet memiliki 2-9
floret yang susunannya mirip dengan spikelet, namun lebih padat. Bagian paling
bawah spikelet adalah dua buah sekam. Sekam pada bagian paling bawah
disebut lower glume, sedangkan sekam yang berada di atasnya disebut upper
glume, keduanya tersusun berhadapan, sehingga menutup sejumlah bunga
tunggal yang ada pada spikelet. Sekam umumnya memiliki struktur keras, pada
saat masih muda berwarna hijau hingga keunguan, dan pada saat masak
fisiologis berwarna putih, cokelat, merah atau hitam, bergantung pada varietas.
Pada ujung spikelet juga sering terdapat 1-2 bunga tunggal yang abnormal akibat
tidak sempurnanya perkembangan benang sari dan bakal biji (Percival 1921,
Allan 1980, Australian Government 2008, Kirby 2002, Anderson and Garlinge
2000).
Bunga tunggal (floret) gandum terbungkus oleh dua buah sekam yang
disebut lemma dan palea. Lemma adalah sekam paling luar dari bunga tunggal,
ujung dari lemma umumnya memiliki bulu (awn). Ukuran bulu pada lemma
beragam, mulai sangat panjang hingga sangat pendek bahkan gundil (tanpa
bulu), bergantung pada varietas. Bulu yang terdapat pada bagian tengah malai
umumnya paling panjang di antara bulu pada bagian yang lain. Warna bulu
bervariasi, mulai dari putih, merah hingga hitam. Bentuk bulu juga beragam
dari lurus hingga seperti spiral (Percival 1921, Allan 1980, Kirby 2002, Anderson
and Garlinge 2000). Pada gandum yang ditanam di daerah kering atau tercekam
kekeringan, buluh berperan dalam membantu proses fotosintesis, bahkan
mampu menyumbang 10-20% bobot kering biji, karena daun mengalami
pengeringan (scenescence) dengan cepat. Pada kondisi suhu udara yang rendah
dan curah hujan tinggi, pengaruh bulu terhadap fotosintesis tidak nyata (Paulsen
1997). Palea merupakan sekam yang terletak di atas lemma, keduanya
menyelimuti bagian reproduksi bunga. Umumnya palea memiliki tekstur yang
lebih lembut dari lemma (Percival 1921, Allan 1980, Kirby 2002, Anderson and
Garlinge 2000).
Dalam setiap bunga tunggal yang normal terdapat tiga benang sari (filament)
dan satu putik (stigma). Benang sari gandum memiliki tangkai sari (stamen)
yang melekatkannya pada dasar bunga. Pada saat masih muda, tangkai sari
berukuran 2-3 mm, namun tumbuh dengan cepat pada saat kepala sari mulai
masak, pada saat bunga mekar ukurannya menjadi 7-10 mm, sehingga kotak
sari akan terjuntai keluar dari lemma dan palea. Benang sari pada gandum
masing-masing berukuran panjang 3-4 mm dan lebar 0.5-1 mm, memiliki dua
kotak sari (locule) berbetuk bi-lobed (dua tabung kembar). Pada masing kotak
sari terdapat 1.000-3.800 serbuk sari (pollen). Pollen yang telah masak (mature)
akan mendesak kotak sari dan keluar dari kotak sari melalui lubang (pore).
Pollen berbentuk bulat, agak oval atau bahkan tidak beraturan dengan diameter
57-65 μ. Putik bunga gandum tersusun atas kepala putik yang bercabang dan
berbulu, dan ovarium (ovary). Ovarium pada bunga gandum terdapat dalam
dua lodikula (lodicules) (Percival 1921, de Vreis 1971, Allan 1980, Australian
Government 2008, Kirby 2002, Anderson and Garlinge 2000).
Biji
Bagian tanaman gandum yang memiliki nilai ekonomi tinggi adalah bijinya. Bagi
tanaman gandum, biji merupakan alat perkembangbiakan, karena dalam biji
terdapat embrio (embryo) yang akan tumbuh menjadi tanaman yang baru. Biji
gandum bekeping satu dan keras sehingga sering disebut kariopsis (caryopsis).
Jumlah biji yang terbentuk dalam setiap spike bila semua bakal biji terserbuki
dan tumbuh normal mencapai lima biji/spike, sedangkan jumlah spike bisa
mencapai 20 spike/malai, namun jumlah biji per malai 10-60 biji. Panjang biji 3-
8 mm, dengan bobot 1.000 biji berkisar antara 15-44 g. Bobot biji gandum akan
menurun bila suhu udara dan suhu tanah meningkat. Dengan bobot 1.000 biji
15-44 g, kebutuhan benih dalam 1 ha sekitar 50 kg (Percival 1921, Kirby 2002,
Hossain et al. 2013, Wahyu et al. 2013, Wirawan et al. 2013). Biji gandum pada
sisi belakang (dorsal) membulat dan halus, sedangkan bagian depan (ventral)
terdapat lekuk (crease), dan pada bagian ujung terdapat rambut halus. Embrio
terdapat pada bagian pangkal biji sisi bagian atas yang tertutup oleh lapisan
skutellum (scutellum) (Gambar 9).
Biji gandum terdiri atas embrio dan skutellum (germ), 3% dari bobot biji,
14% lapisan dedak/bekatul (bran), dan yang paling banyak adalah endosperma
(endosperm), 83% dari total bobot biji (Percival 1921, Kirby 2002, Australian
Government 2008) (Gambar 10).
Pada bagian terluar dari biji gandum terdapat lapisan dedak/bekatul (bran)
yang tersusun atas tiga lapisan yaitu 1) pericarp, 2) testa dan 3) aleuron (Percival
1921, Anderson and Garlinge 2000, Australian Government 2008) (Gambar 11).
Pericarp merupakan bagian terluar dari lapisan dedak, berupa lapisan dengan
ketebalan 45-50 μ, yang tersusun atas 4-5 lapisan sel, yaitu satu lapis sel epidermis
luar, tiga lapis sel parenkim, dan satu lapis sel epidermis dalam. Di bawah pericarp
terdapat testa yang merupakan dua lapis sel panjang, sel pertama tidak berwarna
sedang sel berikutnya memiliki warna, sehingga biji yang masak akan berwarna
putih, krem, cokelat, merah atau hitam. Aleuron merupakan lapisan antara dedak
dengan endosperm, pada aleuron terdapat lapisan lilin dan minyak (Percival
1921, Anderson and Garlinge 2000).
Endosperm merupakan bagian yang paling banyak dari biji gandum,
tersusun atas pati (starch), protein, dan glutein (gluten). Dalam pati/tepung
gandum terdapat banyak vitamin dan mineral yang bermanfaat bagi
pertumbuhan kecambah dan bagi manusia sebagai bahan pangan (Percival
1921, Anderson and Garlinge 2000) (Gambar 12).
Embrio yang terlindungi oleh skutelum disebut germ atau mata tunas.
Skutelum yang menyelimuti embrio memiliki ketebalan 2,5-3 mm, memisahkan
embrio dari endosperma, merupakan jaringan yang termodifikasi dari kotiledon
pada tanaman biji berkeping dua. Skutelum berfungsi pula sebagai tempat
sekresi enzim-enzim yang dibutuhkan biji saat berkecambah dan mengabsorbsi
gula dari hasil pemecahan pati pada endosperm. Embrio gandum tersusun
atas bakal tunas (plumule) dan bakal akar (radicule) (Percival 1921, Anderson
and Garlinge 2000, Kirby 2002) (Gambar 13). Bakal tunas gandum sudah memiliki
empat primordial daun dan primordial tunas (primordial shoot apex). Bakal
akar juga sudah memiliki primordial akar primer (radicle) dan primordial akar
seminal (seminal root) (Percival 1921, Kirby 2002)
tahap pertumbuhan tanaman gandum sama dengan tahap pertumbuhan serealia
berbiji kecil lainnya seperti oat, barley, rye, dan padi (Large 1945, Zadoks et al.
1974). Waktu yang dibutuhkan dalam setiap tahap pertumbuhan bergantung
pada varietas, unsur hara, suhu dan kelembaban, serta dipengaruhi oleh hama
dan penyakit. Terdapat beberapa teori yang menjelaskan tahap -tahap pertumbuhan
tanaman gandum, mulai dari perkecambahan hingga masak fisiologis. Beberapa
teori tentang tahap pertumbuhan gandum antara lain yang dikemukakan oleh
Feekes (1941), Haun (1973), dan Zadoks (1974).
Feekes (1941) membagi tahap -tahap pertumbuhan gandum berdasarkan
perubahan fisik tanaman, mulai dari muncul daum pertama hingga masak
fisiologis. Teori tahap pertumbuhan ini umumnya digunakan membantu dalam
pengendalian hama, penyakit dan gulma (Large 1954, Simmons et al. 1995).
tahap pertumbuhan ini sering digunakan tetapi kurang populer karena kurang
terperinci (Simmons et al. 1995).
tahap pertumbuhan gandum yang dikembangkan Haun (1993) berdasarkan
pertumbuhan atau jumlah daun. Satu unit tahap pertumbuhan ditandai oleh
pertumbuhan atau pertambahan daun baru. tahap pertumbuhan ini juga kurang
terinci dalam menjelaskan tahap pertumbuhan anakan dan tahap perkembangan
biji, sehingga kurang praktis untuk pengamatan di lapangan, karena diperlukan
ketelitian dalam menetukan status dari tahap pertumbuhan yang sedang diamati
(Haun 1973, Simmons et al. 1995).
Teori tahap pertumbuhan gandum yang paling banyak digunakan dalam
menentukan tahap pertumbuhan gandum karena mampu menjelaskan
pertumbuhan gandum secara lebih terinci dan mudah diaplikasikan di lapangan
adalah yang diuraikan oleh Zadoks (1974). tahap pertumbuhan ini menggunakan
dua digit angka, digit pertama menjelaskan tahap pertumbuhan utama,
sedangkan digit kedua menjelaskan tahap pertumbuhan yang sedang terjadi
(Zadoks et al. 1974, Simmons et al. 1995).
tahap pertumbuhan gandum dikembangkan oleh Feekes (1941), Haun
(1973), dan Zadoks (1974), masing-masing memiliki skala yang berbeda. Feekes
(1941) membagi skala pertumbuhan gandum dalam 11 tahap , Haun (1973) 15
tahap utama dan Zadoks (1974) 10 tahap utama. Berdasarkan dari ketiga teori
ini maka secara umum tahap pertumbuhan gandum terbagi dalam 10 tahap
utama, yaitu tahap 0=perkecambahan, tahap 1=pertumbuhan kecambah, tahap
2=pertumbuhan anakan, tahap 3=perpanjangan batang, tahap 4=bunting, tahap
5=pembungaan, tahap 6=antesis, tahap 7=masak susu, tahap 8=masak
adonan,dan tahap 9=pemasakan biji (Gambar 14).
tahap Perkecambahan
tahap perkecambahan biji gandum (Tabel 1), diawali dengan proses imbibisi air
dan oksigen oleh biji. Tahap ini sangat bergantung pada kelembaban tanah dan
suhu lingkungan tumbuh, sehingga persiapan lahan sangat penting (Paulsen
1997, Anderson and Garlinge 2000, Australian Government 2008, CIMMYT 2014).
Air dan oksigen dibutuhkan untuk memecah karbohidrat menjadi gula, yang
dibutuhkan oleh primordial akar dan tunas untuk tumbuh sebelum mampu
mendapatkan nutrisi dari lingkungan tumbuhnya (Anderson and Garlinge 2000).
Hama utama pada tahap ini adalah tikus dan burung yang memakan biji (CIMMYT
2014).
Tahap setelah imbibisi adalah tahap perkecambahan biji yang masih berada
dalam tanah. Bagian pertama muncul yang dari biji berkecambah adalah radikula
(radicle), akan tumbuh pertama, kemudian diikuti oleh koleoptil (coleoptiles)
(Gambar 15a). Pada umumnya koleoptil akan muncul ke permukaan tanah
dalam waktu 1-2 hari, bergantung pada kedalaman penanaman biji dan kondisi
lingkungan. Munculnya koleoptil ke permukaan tanah disebut seed emergence.
Kecambah muncul pada permukaan tanah 5-7 hari bila kondisi lingkungan
kurang mendukung dan ditanam terlalu dalam. Kecambah yang tumbuh 4-5
hari umumnya kurang toleran cekaman kekeringan (Paulsen 1997, Anderson
and Garlinge 2000, Australian Government 2008, Nur et al. 2010).
tahap Pertumbuhan Kecambah (Seedling Growth)
Setelah koleoptil muncul ke permukaan tanah maka pertumbuhannya akan
terhenti dan dari ujung koleoptil akan muncul daun pertama yang tumbuh dari
titik tumbuh (growing point) yang masih berada di bawah permukaan tanah
(Gambar 16a). Koleoptil berfungsi melindungi daun pertama selama proses
menembus permukaan tanah. tahap pertumbuhan kecambah berlangsung
hingga terbentuk 5-9 buah daun, bergantung pada varietas dan kondisi
lingkungan (Gambar 16b) (Tabel 2). Varietas berumur dalam memiliki daun
yang lebih banyak. Sebagai contoh, gandum musim dingin yang berumur dalam
memiliki 11-15 daun. Pada tahap ini penyiangan sangat penting karena persaingan
dengan gulma masih tinggi dan titik tumbuh masih di bawah permukaan tanah
(Paulsen 1997, Simmons et al. 1995, Acevedo et al. 2002, Anderson and Garlinge
2000, Australian Government 2008, CIMMYT 2014)
tahap Pertumbuhan Anakan (Tillering)
Anakan adalah tunas yang tumbuh dari tunas buku (nodal bud) pada batang
utama. Pada tahap ini titik tumbuh juga masih di bawah permukaan tanah.
Pertumbuhan anakan erat kaitanya dengan laju pertumbuhan daun. Anakan
pertama tumbuh pada filokron pertama, yaitu di antara koleoptil dan daun
pertama (Gambar 17), pada saat tanaman berumur kurang lebih 30 HST (hari
setelah tanam). Filokron (phyllochronus) merupakan jarak antara daun dan
daun berikutnya. Jumlah anakan bergantung pada varietas dan kondisi
lingkungan tumbuh. Pertumbuhan daun pada anakan memiliki pola yang sama
dengan batang utama. Anakan yang tumbuh mendekati tahap pembungaan akan
mati (scenescence) terlebih dahulu. Anakan sangat penting bagi tanaman
gandum sebagai kompensasi bila jarak tanam terlalu lebar atau bila batang
utama gagal berkembang akibat cekaman lingkungan, baik biotik maupun
abiotik. Namun cekaman kekeringan atau suhu yang rendah pada tahap ini akan
menurunkan jumlah anakan. Pada tahap ini penyiangan dan pemupukan
diperlukan untuk pertumbuhan optimal, terutama pupuk N (Paulsen 1997,
Simmons et al. 1995, Anderson and Garlinge 2000, Australian Government 2008,
Nur et al. 2010, CIMMYT 2014).
tahap Perpanjangan Batang (Stem elongation/Jointing)
tahap perpanjangan batang merupakan proses memanjangnya ruas-ruas pada
batang gandum. tahap ini berkaitan dengan laju pertumbuhan daun, anakan,
akar dan pembungaan. tahap ini dimulai pada saat jumlah anakan sudah cukup
dan primordial bunga sudah berkembang sempurna, dimana benang sari sudah
tumbuh dan spikelet terakhir (terminal spikelet) juga sudah terbentuk, meski
dalam ukuran yang masih sangat kecil tersimpan pada titik tumbuh (Gambar
18 dan 19). Spikelet terakhir terbentuk pada saat tanaman memiliki 7-12 daun.
Pada tahap ini umumnya batang utama telah memiliki setidaknya lima daun. tahap
pemanjangan batang akan berakhir pada saat daun bendera benar-benar
terbuka, yang ditandai oleh terlihatnya ligule daun bendera (Tabel 4), namun
pertambahan tinggi tanaman masih terus berlangsung hingga tanaman
memasuki tahap antesis (anthesis) (Gambar 20). Bakal malai tumbuh sebelum
daun bendera muncul, hingga 10 hari setelah antesis. Pada saat daun bendera
belum muncul, pertumbuhan malai lambat. Pada saat ligula daun bendera
muncul, pertumbuhan sangat cepat (Paulsen 1997, Simmons et al. 1995,
Anderson and Garlinge 2000, Acevedo et al. 2002, Australian Government 2008).
Pertumbuhan batang mendorong titik tumbuh ke atas permukaan tanah.
Ruas yang pertama kali memanjang adalah ruas ke-4 dari bawah (Gambar 21).
Pada saat ruas ini memanjang hingga 50% dari ukuran panjangnya, maka ruas
di atasnya mulai memanjang (Gambar 22), begitu seterusnya hingga ruas terakhir
(peduncle), yang membawa malai. Peduncle merupakan ruas yang paling
panjang pada tanaman gandum. Panjang/tinggi tanaman gandum beragam
dari 30-150 cm, bergantung pada varietas dan lingkungan tumbuh. Tinggi
tanaman lebih disebabkan oleh ukuran ruas dibandingkan jumlah buku. Tanaman
gandum berbatang pendek umumnya lebih toleran rebah. Pada tahap ini hara
dan air yang cukup sangat penting karena jumlah spikelet dan floret mulai
berkembang dan merupakan tahap pertumbuhan yang paling cepat, begitu juga
laju penyerapan unsur hara yang sangat tinggi, sehingga kekurangan hara dan
cekaman lingkungan akan menurunkan hasil, terutama hara N dan air (Paulsen
1997, Simmons et al. 1995, Anderson and Garlinge 2000, Acevedo et al. 2002,
Australian Government 2008, CIMMYT 2014).
tahap Bunting (Booting)
tahap bunting tanaman gandum merupakan awal dari tahap reproduktif meskipun
sebenarnya telah dimulai sebelum tanaman memasuki tahap pemanjangan
batang. tahap ini ditandai oleh pemanjangan pelepah daun bendera dan diakhiri
oleh munculnya buluh (Gambar 23) (Tabel 5). Pertumbuhan malai pada pelepah
daun mulai cepat pada saat ligula daun bendera terlihat. Pelepah daun bendera
semakin membengkak, akibat dorongan pertumbuhan malai di dalamnya. Pada
tahap ini cekaman kekeringan, suhu udara yang rendah, dan penyinaran yang
terlalu tinggi atau terlalu rendah berpengaruh terhadap jumlah malai pada saat
panen, karena pertumbuhan bunga fertile dimulai pada tahap ini. Suhu udara
yang terlalu rendah atau terlalu tinggi memicu banyak bunga tunggal
(floret) yang steril (Simmons et al. 1995, Warrington et al. 1977, Anderson and
Garlinge 2000, Acevedo et al. 2002, CIMMYT 2014).
tahap Pembungaan (Infloresence Emergence/Heading)
tahap pembungaan tanaman gandum diawali oleh munculnya malai dari dalam
pelepah daun bendera (Tabel 6) (Gambar 24). Umur berbunga gandum berbeda
dengan jagung. Pada jagung, umur berbunga ditentukan jika 50% kotak sari
pada poros utama malai telah pecah. Pada gandum, sorgum, padi, oat barley
dan rye, umur berbunga adalah jika malai telah 50% keluar dari dalam pelepah
daun bendera. Pada tahap ini pertumbuhan malai dalam pelepah daun bendera
masih terus berlangsung dan semakin cepat, hingga mendekati antesis. Proses
perpanjangan ruas terakhir (peduncle) juga masih terus berlangsung. Di dalam
malai, bunga tunggal juga terus berkembang, beberapa bunga tunggal akan
degenerasi/luruh/aborsi akibat persaingan nutrisi antarbunga tunggal (Gambar
25). Bunga yang mengalami degenerasi ada yang luruh/aborsi, atau akan menjadi
bunga abnormal. Umumnya bunga abnormal masih memiliki anter tetapi bakal
bijinya tidak berkembang. Bunga yang mengalami degenerasi umumnya bunga
tunggal yang berada paling jauh dari tangkai malai utama, seperti bunga tunggal
yang berada paling ujung (terminal spikelet/floret) dan bunga tunggal yang
berada pada pangkal tangkai utama (Gambar 26). Munculnya malai pada batang
utama umumnya lebih awal dibanding anakan. tahap ini rentan terhadap
kekurangan air. Umur berbunga gandum yang ditanam pada ketinggian di atas
1.000 mdpl antara berkisar 43-65 HST, sedangkan di dataran rendah ≤ 500 mdpl
berkisar antara 43-65 HST (Paulsen 1997, Simmons et al. 1995, Acevedo et al.
2002, Nur et al. 2010, Wirawan et al. 2013, Wahyu et al. 2013, CIMMYT 2014).
tahap Antesis (Anthesis)
tahap antesis merupakan tahap pecahnya kotak sari (anther) dan tersebarnya serbuk
sari keluar kotak sari. Bunga gandum termasuk klestogami, yaitu tanaman
menyerbuk sendiri (self pollination), di Maros Sulawesi Selatan, kotak sari telah
pecah sebelum keluar dari sekam. Pada saat kotak sari keluar dari sekam (lemma
dan palea) merupakan pertanda proses penyerbukan sudah terjadi, 5-7% serbuk
sari sudah tersebar di dalam sekam. Kotak sari yang keluar dari sekam berwarna
kuning hingga putih (de Vreis 1971, Allan 1980, Paulsen 1997, Simmons et al. 1995,
Anderson and Garlinge 2000, Acevedo et al. 2002, Australian Government 2008).
tahap antesis diawali dengan pecahnya sebuk sari dan diakhiri munculnya
kotak sari di seluruh bagian malai (Gambar 27) (Tabel 7). Kotak sari yang pertama
pecah adalah kotak sari pada bunga tunggal (floret) yang terletak pada 1/2
bagian tengah malai, kemudian bagian atas malai dan terakhir adalah bunga
tunggal yang terletak pada bagian paling bawah malai. Proses antesis dari awal
hingga malai 100% antesis memerlukan waktu 3-5 hari. Bunga akan mekar setiap
waktu pada siang hari, namun kebanyakan terjadi pada pagi hari dan hanya
sedikit yang mekar pada siang hari, bergantung pada varietas dan kondisi
lingkungan. Bunga tunggal gandum akan mekar hanya dalam waktu 8-60 menit.
Putik (stigma) akan terus reseptif (receptive) hingga 13 hari setelah antesis,
namun yang paling reseptif adalah 0-3 hari setelah antesis. Viabilitas serbuk sari
di udara bebas kurang dari 30 menit (Percival 1921, de Vreis 1971, Allan 1980,
Paulsen 1997, Simmons et al. 1995, Anderson and Garlinge 2000, Acevedo et al.
2002, Australian Government 2008).
Peluang gandum untuk menyerbuk silang adalah ± 10%, bergantung pada
kepadatan populasi dan kondisi lingkungan. Pada kondisi cuaca yang kering
dan hangat, peluang menyerbuk silang 3,7-9,7%. Pada kondisi kelembaban
udara yang tinggi, peluang menyerbuk silangnya 0,1% dan pada musim dingin
0,1-5,6%. Lebih dari 90% serbuk sari gandum jatuh pada radius 3 m dari tanaman
(Acevedo et al. 2002, Nevo et al. 2002, Australian Government 2008). Cekaman
kekurangan air 7 hari menjelang dan saat antesis dapat menurunkan hasil biji
per malai (Wardlaw 1971).
tahap Masak Susu (Milk development)
Setelah terjadi proses penyerbukan maka proses pembuahan akan segera
dimulai, diawali dengan perkecambahan serbuk sari di atas putik, proses ini
terjadi hanya beberapa menit setelah serbuk sari mendarat di atas permukaan
putik yang reseptif. Serbuk sari yang berkecambah akan membentuk tabung
sari (pollen tube). Tabung sari tumbuh 1-2 jam setelah penyerbukan. Proses
selanjutnya adalah pembuahan yang terjadi 30-40 jam setelah penyerbukan.
(Percival 1921, de Vreis 1971, Australian Government 2008).
Setelah terjadi proses pembuahan maka endosperm (bakal biji/kernel) dan
embrio pada bakal biji segera berkembang. Laju pertumbuhan biji dan embrio
bergantung pada varietas dan lingkungan. Ukuran biji bertambah dengan cepat,
7-14 hari setelah penyerbukan ukuran biji sudah mencapai maksimum, tetapi
penimbunan pati belum maksimal, begitu pula pertumbuhan embrio. Ukuran
biji muda akan tetap hingga masak fisiologis (Gambar 28 dan 29). (Percival 1921,
Paulsen 1997, Simmons et al. 1995, Australian Government 2008, Wheat BP 2014).
Setelah ukuran endosperm mencapai maksimum, akumulasi pati dan
protein segera meningkat dengan cepat. Pada awal akumulasi pati dan protein,
endosperm masih berupa cairan, tahap ini disebut tahap masak susu. tahap masak
susu diawali pada saat cairan endosperm masih berupa air hingga berupa cairan
yang menyerupai susu (Tabel 8) (Percival 1921, Paulsen 1997, Simmons et al.
1995, Australian Government 2008, Herbek and Lee 2009). Pada tahap masak
susu ini, sekam dan biji masih berwarna hijau, embrio masih dapat dipisahkan
dengan mudah dari endosperm, dan jika ditekan biji akan mudah pecah dan
berisi cairan seperti susu (Gambar 30). tahap paling awal dari tahap masak susu
adalah tahap masak air (water ripe), yaitu endosperm hanya berisi air. Pada tahap
ini embrio masih sangat kecil, berada pada tahap globular. tahap masak air terjadi
4-10 hari setelah pembuahan. tahap selanjutnya adalah masak susu awal (early
milk), yaitu cairan dalam endosperm tampak seperti susu. Hal ini terjadi karena
butiran pati telah terbentuk. Pada tahap ini ukuran embrio bertambah tapi masih
kecil. Peningkatan kandungan pati memicu kekentalan cairan dalam
endosperm bertambah, pada 11-16 hari setelah pembuahan biji memasuki tahap
masak susu pertengahan (medium milk). Pada tahap ini embrio tumbuh dengan
cepat. Pada hari ke-16, biji memasuki tahap masak susu akhir (late milk), pada
tahap cairan seperti susu yang kental, embrio telah terbentuk sempurna begitu
pula dengan skutelum (Percival 1921, Paulsen 1997, Simmons et al. 1995,
Australian Government 2008, Herbek and Lee 2009).
tahap Masak Adonan (Dough stage)
tahap masak adonan merupakan tahap akumulasi pati pada endosperm meningkat
dengan sangat cepat. Cairan dalam endosperm biji semakin mengental jika
ditekan sehingga seperti adonan kue. Oleh karena itu, tahap ini disebut sebagai
tahap masak adonan. Pada tahap ini, sekam dan biji akan mulai berubah warna
menjadi kekuningan atau pucat (Gambar 31) (Tabel 9). tahap masak adonan
diawali dengan tahap awal masak adonan (early dough) (Gambar 32a). Pada tahap
ini, akumulasi kandungan air masih banyak tetapi sudah mulai mengental. tahap
selanjutnya adalah tahap masak adonan lunak (soft dough) (Gambar 32b). Pada
tahap ini, endosperm terasa lembut tetapi kering karena kandungan air sudah
turun. Akumulasi bahan kering biji sebagian besar sudah tercapai. Embrio sudah
hampir mendekati ukuran maksimum. tahap masak adonan terjadi antara 17-21
hari setelah penyerbukan. tahap terakhir dari tahap ini adalah masak adonan keras
(hard dough) (Gambar 32c). Pada tahap ini, endosperm sudah keras jika ditekan
dengan jari, biji sudah tidak bisa pecah tetapi meninggalkan bekas kuku (dented).
Kandungan air pada endosperm berkisar antara 30-40%. Bobot kering kernel
sudah mencapai masimum. tahap ini terjadi pada 21-30 hari setelah penyerbukan.
Pada tahap masak adonan, tanaman rentan terhadap rebah (Percival 1921, Paulsen
1997, Simmons et al. 1995, Australian Government 2008, Herbek and Lee 2009).
tahap Pemasakan Biji (Ripening)
tahap pemasakan gandum terjadi saat biji berumur 30-40 hari setelah
penyerbukan, bergantung pada varietas dan kondisi lingkungan. Umur masak
fisiologis gandum yang ditanam di dataran tinggi ≥1000 mdpl berkisar antara
94-105 HST, sedangkan yang ditanam pada ketinggian ≤ 500 mdpl lebih cepat,
73-93 HST. Pada tahap ini tanaman mulai mengering, warna biji berubah semakin
kuning tua/cokelat/merah, bergantung pada varietas (Tabel 10). Biji pada tahap
ini terasa keras saat ditekan dengan kuku, dan sulit pecah (Gambar 33).
Kandungan air biji akan terus turun, hingga saat masak fisiologis kandungan air
biji 13-12%, dan tanaman siap dipanen. Memasuki umur panen, biji gandum
yang kering adakalanya tampak keluar dari sekam, bahkan pada varietas yang
mudah rontok, biji akan rontok di lapang bila kondisi sangat kering. Namun bila
kondisi udara lembab, biji yang sudah masuk masa panen akan tumbuh di
malai (Gambar 34). Biji gandum sebaiknya dipanen pada kadar air biji kurang
dari 20%, untuk mencegah kerusakan biji (Percival 1921, Paulsen 1997, Simmons
et al. 1995, Australian Government 2008, Herbek and Lee 2009, Nur et al. 2010,
Wirawan et al. 2013, Wahyu et al. 2013).
Gandum (Triticum tcc.) merupakan tanaman serealia penting ketiga di dunia
setelah padi dan jagung. Tanaman C3 ini dapat beradaptasi baik pada daerah
dingin. Suhu optimum untuk pertumbuhan gandum berkisar antara 20-25°C,
sedangkan suhu tanah minimum yang dibutuhkan adalah 2-4°C. Secara
agronomis, gandum di negara kita dapat tumbuh dan berproduksi dengan baik
pada lokasi dengan ketinggian > 1.000 di atas permukaan laut (dpl), atau dataran
rendah dengan sifat lahan beriklim kering dan suhu < 25°C seperti di
sebagian wilayah NTT (Balitsereal 2012). Potensi lahan untuk pengembangan
gandum di negara kita cukup besar, khususnya pada lahan dataran tinggi, untuk
memutus siklus penyebaran hama penyakit tanaman hortikultura seperti
kentang atau tomat. Pada wilayah ini gandum dapat tumbuh baik bila
penanamannya disesuaikan dengan kondisi kelembaban dan curah hujan.
Kebutuhan air bagi tanaman gandum untuk berproduksi optimal berkisar
antara 450-650 mm, bergantung pada iklim dan lama pertumbuhan tanaman
(FAO 2001). Di Asia, khususnya Asia Selatan, kebutuhan air bagi tanaman gandum
cenderung lebih sedikit, berkisar antara 400-450 mm (Ministry of Agriculture,
India 2014). Hal ini disebabkan oleh umur tanaman yang relatif lebih pendek. Di
negara kita dengan kondisi iklim yang lembab dan suhu tinggi, tanaman gandum
dapat dipanen pada umur 85-115 hari, bergantung pada varietas dan suhu
lingkungan tumbuh (Aqil dan Rapar 2013). Ketepatan ketersediaan air pada
stadia pertumbuhan berpengaruh terhadap produksi gandum. Hasil optimal
akan tercapai apabila tanaman mendapat pengairan pada tahap pembentukan
rumpun, pembungaan, dan pengisian bulir. Oleh karena itu diperlukan teknologi
pengelolaan air yang tepat sesuai kebutuhan tanaman gandum.
Pengelolaan air perlu disesuaikan dengan sumber daya fisik alam (tanah,
iklim, sumber air) dan biologi dengan memanfaatkan berbagai disiplin ilmu
untuk membawa air ke perakaran tanaman sehingga mampu meningkatkan
produksi (Nobe and Sampath 1986). Sasaran dari pengelolaan air adalah
tercapainya empat tujuan pokok, yaitu: (1) efisiensi pemakaian air dan produksi
tanaman yang tinggi; (2) efisiensi biaya pemakaian air; (3) pemerataan
pemakaian air yang terbatas, baik dari segi waktu maupun jumlah; dan (4)
keberlanjutan sistem pemakaian sumber daya air. Dalam hubungannya
dengan pengelolaan air untuk tanaman gandum yang banyak dibudidayakan
di dataran tinggi, pengelolaan agroklimat penting untuk diperhatikan. Tulisan
ini membahas pengelolaan air untuk tanaman gandum yang mencakup
sifat wilayah dan pola tanam, kebutuhan air tanaman, hubungan tingkat
pemberian air dengan tingkat hasil, sistem perakaran dan metode pemberian
air bagi tanaman.
sifat WILAYAH DAN POLA TANAM GANDUM
DI negara kita
Pertanaman gandum di negara kita umumnya dijumpai pada wilayah dataran
tinggi (> 1.000 m dpl) atau pada dataran rendah dengan suhu dan kelembaban
yang rendah (<25o
C). Faktor kelembaban juga penting dalam budi daya gandum
untuk menekan perkembangan penyakit/jamur. Gandum juga tidak sesuai
ditanam pada wilayah dengan curah hujan tinggi. Curah hujan optimum untuk
pertumbuhan gandum berkisar antara 350-1.250 mm. Gandum umumnya
tumbuh baik pada tanah Andosol, Regosol, Latosol dan Alluvial dengan pH 6-7.
Tanah yang baik untuk pertumbuhan gandum adalah bertekstur sedang/
medium. Tanah gambut atau tanah dengan kandungan S, Mg atau Fe yang
tinggi kurang sesuai untuk budi daya gandum. Wilayah penghasil gandum di
negara kita di antaranya Pengalengan, Dieng, Tengger, Karanganyar, Kopeng
Salaran, Piji Salatiga Jawa Tengah, Malino, Enrekang dan Bantaeng Sulawesi
Selatan, Tomohon Sulawesi Utara, Napu Sulawesi Tengah, Merauke Papua, dan
NTT (Balitsereal 2013).
sifat lahan yang sesuai untuk tanaman gandum dapat dilihat pada
Tabel 1. Parameter yang digunakan dalam menilai kesesuaian lahan meliputi
temperatur, curah hujan, kelembaban udara, drainase, tekstur, bahan kasar,
kedalaman tanah, ketebalan gambut, kapasitas tukar kation liat, kejenuhan basa,
pH H2O, C-organik, salinitas, alkalinitas, kedalaman bahan sulfidik, lereng,
bahaya erosi, dan genangan.
Berdasarkan sifat kualitas lahan yang ditunjukkan pada Tabel 1,
kelas kesesuaian lahan untuk tanaman gandum dibedakan menjadi S1 (sangat
sesuai), yaitu apabila lahan tidak mempunyai faktor pembatas yang berarti atau
faktor pembatas bersifat minor dan tidak akan mereduksi produktvitas lahan
secara nyata; S2 (cukup sesuai), yaitu lahan dengan faktor pembatas yang
mempengaruhi produktivitas tanaman secara nyata dan memerlukan tambahan
masukkan atau input; S3 (sesuai marjinal), yaitu lahan dengan faktor pembatas
yang berat yang nyata mempengaruhi produktivitas. Faktor pembatas
pemakaian lahan kelas S2 dapat diatasi oleh pelaku pertanian atau petani,
sedangkan lahan kelas S3 memerlukan tambahan input yang relatif lebih banyak
dibandingkan dengan kelas S2. Lahan kelas N tidak sesuai untuk budi daya
gandum karena memiliki faktor pembatas yang lebih berat (Djaenudin et al.
2003).
Pengembangan gandum di negara kita saat ini masih menghadapi sejumlah
kendala, terutama terbatasnya luas lahan untuk pengembangan dan kompetisi
dengan tanaman hortikultura seperti kentang atau tomat. Farid (2006)
menyatakan bahwa terbatasnya luas lahan di dataran tinggi yang banyak
ditanami dengan komoditas hortikultura yang mempunyai nilai ekonomis tinggi
juga berpengaruh terhadap pengembangan gandum di negara kita , sehingga
diperlukan varietas gandum yang dapat beradaptasi di dataran rendah (< 400
m dpl). Kendala utama yang dihadapi dalam pengembangan gandum di dataran
rendah adalah cekaman lingkungan, khususnya suhu tinggi.
Gandum dapat dibudidayakan secara monokultur atau pergiliran dengan
tanaman hortikultura untuk memutus siklus hama penyakit. Berdasarkan
peluang dan kejadian hujan, gandum dapat dibudidayakan dengan pola rotasi
sebagai berikut:
Dataran tinggi (>1.000 m dpl)
Kentang – gandum – tomat
Kentang – wortel – gandum
Kentang – gandum – bera
Dataran menengah (600-800 m dpl)
Gandum – gandum – wortel
Tomat – gandum – bera
Dataran rendah (suhu 20-25o
C)
Padi – gandum – bera
Hortikultura – gandum – bera
Padi – gandum – hortikultura
KEBUTUHAN AIR TANAMAN
Pola curah hujan secara umum digunakan sebagai acuan dalam menentukan
kecukupan air bagi tanaman gandum. Pola hujan juga dijadikan acuan dalam
perencanaan pengairan tanaman. Dalam kaitannya dengan perhitungan
matematis kecukupan air tanaman, parameter yang perlu mendapat perhatian
adalah evapotranspirasi.
Evapotranspirasi adalah kombinasi proses kehilangan air dari suatu lahan
melalui evaporasi dan transpirasi. Evaporasi adalah proses dimana air diubah
menjadi uap air dan selanjutnya uap air dipindahkan dari permukaan bidang
penguapan ke atmosfer. Transpirasi adalah vaporisasi dalam jaringan tanaman
dan selanjutnya uap air dipindahkan dari permukaan tanaman ke atmosfer
(vapor removal). Pada transpirasi, vaporisasi terjadi terutama di ruang antarsel
daun dan selanjutnya uap air melalui stomata akan lepas ke atmosfer (Allen et
al. 1998). Evapotranspirasi tanaman dapat dikelompokkan menjadi dua bagian,
yaitu evapotranspirasi potensial (ETP) dan evapotranspirasi aktual (ETA).
ETP merupakan jumlah air yang ditranspirasikan dalam satuan unit waktu
oleh tanaman yang menutupi tanah secara keseluruhan dengan ketinggian
seragam, tidak pernah kekurangan air, dan tanaman tidak terserang hama
penyakit. Dengan kata lain, ETP dapat diinterpretasikan sebagai kehilangan air
tanaman yang diakibatkan oleh faktor fisiologis dan klimatologis. Penentuan
nilai kebutuhan air tanaman (evapotranspirasi) sejauh ini masih berdasarkan
pada persamaan empiris yang telah banyak dikembangkan. Di antara
persamaan-persamaan empiris yang umum digunakan adalah metode BlaneyCriddledan, sedangkan pemakaian langsung di lapang umumnya
menggunakan peralatan untuk mengamati perubahan air tanah. ETP dapat
dihitung secara empiris dengan persamaan Penman (Doorenbos and Pruitt
1984) sebagai berikut:
ETP = C (Δ/(Δ + γ) (Rn – G) + γ/ (Δ + γ) 2.7 Wf (ez
o
– ez
))
Dimana:
C = faktor koreksi
Δ = pertambahan tekanan uap jenuh
γ = konstanta psikometrik
Rn = radiasi matahari bersih (mm/hari)
G = fluks panas laten tanah (untuk periode harian = 0)
Wf = fungsi kecepatan angin ( 1 + 0,864 u )
(e z - ez ) = defisit tekanan uap (mbar)
(eo
) = tekanan uap jenuh ( mbar)
() = tekanan uap aktual (mbar)
ETA merupakan tebal air yang dibutuhkan untuk mengganti sejumlah air
yang hilang melalui evapotranspirasi pada tanaman yang sehat. Nilai ETA adalah
nilai kebutuhan air yang harus diberikan ke tanaman, atau merupakan dasar
dalam penentuan kebutuhan air bagi tanaman di lapang dengan persamaan
empiris:
ETA = ETP x Kc
Dimana:
ETA = evapotranspirasi aktual (mm)
ETP = evapotranspirasi potensial (mm)
Kc = koefisien tanaman
Koefisien tanaman (Kc) menggambarkan laju kehilangan air secara drastis
pada tahap -tahap pertumbuhan tanaman dan keseimbangan komponenkomponen energi yang mempengaruhi pertumbuhan tanaman (FAO 2001).
Gambar 1 memperlihatkan tahapan pertumbuhan tanaman gandum dan
koefisien tanaman yang digunakan untuk mengatur pemberian air. Pada awal
pertumbuhan Kc tanaman gandum masih rendah (0,4) sehingga kebutuhan
airnya juga relatif kecil. Namun seiring dengan pertumbuhan, konsumsi air untuk
kegiatan metabolisme tanaman menjadi meningkat dan mencapai puncaknya
pada tahap pembungaan sampai pengisian biji yang ditunjukkan oleh nilai Kc
yang tinggi dan deplesi (evaporasi) air yang juga tinggi.
Penentuan ETA di lapang dapat menggunakan lisimeter, yaitu tangki yang
diisi dengan tanah, ditanami dengan tanaman tertentu, dan diletakkan pada
lahan terbuka. pemakaian tangki evaporasi ini sangat praktis karena hanya
satu parameter saja yang diukur. Parameter ini selanjutnya digabungkan
dengan informasi kebutuhan air tiap musim, umur tanaman gandum, dan luas
lisimeter sehingga kebutuhan air harian atau dekade dapat dihitung dengan
mudah (Doorenbos and Pruitt 1984).
KETERSEDIAAN AIR DAN HUBUNGANNYA DENGAN
HASIL GANDUM
Tanaman gandum dapat tumbuh dengan baik apabila ketersediaan air baik
dalam bentuk hujan maupun irigasi, terjaga selama pertumbuhan berlangsung.
Selain itu faktor ketepatan pemberian air yang sesuai dengan tahap pertumbuhan
tanaman gandum juga menentukan tingkat produksi tanaman. Pada tanaman
gandum, periode pertumbuhan tanaman sesuai dengan tingkat kebutuhan
airnya dibagi menjadi lima tahap , yaitu tahap pertumbuhan awal, tahap vegetatif,
tahap pembungaan, tahap pengisian malai, dan tahap pematangan. tahap
pertumbuhan dan jumlah hari tanaman gandum disajikan pada Tabel 3 dan
Gambar 2.
Setelah melalui tahap pertumbuhan awal (mulai saat tanam sampai menjelang
tahap vegetatif), anakan gandum mulai tumbuh dari axils tunas daun utama.
Potensi jumlah anakan bervariasi dengan genotipe, khususnya di antara jenis
berbunga. Gandum musim dingin (spring wheat) memiliki jumlah anakan yang
lebih besar. Longnecker et al. (1993) menyatakan bahwa anakan terbentuk
pada masa vegetatif awal dan vegetatif akhir, faktor genetik dan lingkungan
sangat berperan dalam proses ini . Selain itu, jumlah anakan produktif
juga dipengaruhi oleh populasi/kerapatan tanaman.
Hubungan tingkat penurunan hasil gandum terhadap defisit
evapotranspirasi relatif selama periode pertumbuhan tanaman disajikan pada
Gambar 4. Table 3 dan Gambar 4 menunjukkan bahwa lama waktu tumbuh
tanaman sampai panen bervariasi antara 100-140 hari. tahap vegetatif sampai
menjelang pembungaan dan tahap pengisian bulir sampai menjelang pemasakan
membutuhkan waktu yang paling lama, berkisar antara 60-70 hari. Hasil biji
juga dipengaruhi oleh lama waktu dan intensitas defisit air serta tahap
pertumbuhan dimana defisit air terjadi. Tanaman gandum musim semi lebih
sensitif terhadap kekurangan air dibandingkan dengan gandum musim dingin.
Hubungan antara tingkat penurunan hasil relatif (1-Ya/Ym) dengan defisit
evapotranspirasi relatif tanaman gandum selama pertumbuhan disajikan pada
Gambar 3. Tanaman gandum lebih toleran terhadap kekurangan air pada tahap
vegetatif akhir (tahap 1) dan tahap pemasakan dibandingkan dengan tahap lainnya.
tahap pembentukan anakan dan pembungaan serta pertumbuhan awal adalah
tahap yang paling kritis dan kekurangan air pada tahap ini akan menimbulkan
kehilangan hasil yang paling tinggi, mencapai 40%.
Kekurangan air pada tahap pembentukan anakan dan pembungaan
memicu pembentukan pollen (serbuk sari) terganggu sehingga jumlah
anakan berkurang. Selain pembentukan anakan terganggu, kekurangan air pada
tahap pembentukan rumpun dan pembungaan juga menurunkan jumlah
rumpun per tanaman, panjang malai, dan jumlah biji per malai. Ottman et al.
(2012) menyatakan bahwa tanaman yang stres kekurangan air pada tahap
penyerbukan mengganggu aktivitas penyerbukan dan mengurangi jumlah biji.
Kekeringan juga mengganggu perkembangan akar bahkan memicu
tanaman menjadi mati. Selain faktor kekurangan air, kondisi cuaca panas yang
disertai angin kering membuat biji menjadi kusut sehingga kualitasnya rendah.
Yang et al. (2000) serta Zhang dan Yang (2004) melaporkan bahwa defisit
lengas tanah pada tahap pengisian biji memicu percepatan penuaan daun
tanaman dan periode pengisian biji menjadi lebih cepat sekitar 10 hari. Hal ini
berdampak kepada percepatan laju pengisian biji dan meningkatnya mobilisasi
cadangan karbohidrat yang berdampak pada penurunan hasil.
Pengaruh kekurangan air pada tahap pertumbuhan awal terhadap hasil biji
dan biomas gandum juga banyak dilaporkan. Kekurangan air pada tahap ini
berpengaruh terhadap akumulasi biomas gandum akibat rendah/menurunnya
indeks luas daun dan efisiensi pemakaian radiasi matahari (Jamieson et al.
1998). Kiniry et al. (1989) melaporkan bahwa intersepsi radiasi matahari
berkorelasi dengan akumulasi bobot kering tanaman gandum.
PENYERAPAN AIR OLEH AKAR TANAMAN GANDUM
Tanaman gandum pada awalnya mempunyai akar primer dan kemudian
berkembang menjadi akar serabut. Akar-akar ini selanjutnya berkembang
dari cabang yang letaknya di sekitar permukaan tanah. Kedalaman dan
kerapatan akar dipengaruhi oleh air, nutrisi, dan ketersediaan oksigen dalam
tanah. Akar tanaman gandum akan berkembang rata-rata sampai 90 cm dan
pada kondisi tertentu mencapai 120 cm. Radius sebaran akar berkisar antara
0,15-0,25 ke semua arah (FAO 2001).
Pola penyerapan dan pemakaian air untuk mendukung pertumbuhan
tanaman dipengaruhi oleh kerapatan akar. Secara umum 50-60% dari total air
diserap akar pada kedalaman 30 cm, sementara 20-25% pada kedalaman akar
30-60 cm, dan 10-15 % sisanya pada kedalaman 60-90 cm. Berdasarkan pola
penyerapan air ini maka pemberian air/irigasi pada tanaman gandum
hendaknya mempertimbangkan pola penyerapan air oleh akar tanaman.
Pengaruh pemberian air dalam jumlah terbatas (limited irrigation) terhadap
pola pembentukan akar dan hasil tanaman gandum telah banyak dilaporkan.
Singh et al. (1991) melaporkan bahwa periode sensitif tanaman terhadap
kecukupan air bervariasi menurut tahapan pertumbuhan tanaman, variabilitas/
keragaman wilayah dan praktek budi daya petani. Pada wilayah subtropis,
respon tanaman gandum terhadap defisit irigasi sangat tinggi pada tahap
pemanjangan batang sampai tahap bunting (Zhang and Oweis 1999).
PENGARUH SUHU/PANAS TERHADAP HASIL GANDUM
Suhu merupakan salah satu faktor pendukung keberhasilan budi daya gandum.
Suhu yang tinggi berdampak pada penurunan hasil akibat terjadinya bulir hampa
(biji tidak terbentuk), memendeknya durasi pembentukan dan pengisian biji,
serta terhambatnya asimilasi sukrosa dalam biji. Asana dan Williams (1965)
dalam penelitiannya pada ruang terkontrol dengan suhu 25-35o
C mendapatkan
bobot biji gandum menurun 16% untuk setiap kenaikan suhu 5oC. Ini berarti
apabila terjadi kenaikan suhu 10
C terjadi penurunan hasil biji sampai 8%,
bertambahnya bulir hampa 5-6%, dan mengecilnya ukuran biji 3-4%. Suhu
lingkungan yang tinggi menghambat proses fisiologi dan biokimia tanaman,
diantaranya terganggunya suplai bahan asimilasi ke biji sehingga berdampak
pada menurunnya akumulasi bobot kering tanaman gandum (Reynolds et al.
2001).
Di antara tahap pertumbuhan tanaman gandum, cekaman suhu tinggi pada
tahap antesis paling besar pengaruhnya terhadap hasil biji karena mempengaruhi
proses penyerbukan dan pengisian biji. Penurunan bobot biji/bulir akibat
pengaruh stres panas berkaitan erat dengan pengaruh suhu terhadap laju dan
lama waktu perkembangan biji/bulir. Peningkatan suhu dari 15/10o
C menjadi
21/16o
C akan mempercepat waktu pengisian biji dari 60 hari menjadi 36 hari
dan laju pertumbuhan biji lebih lebih cepat dari 0,73 mg/biji/hari menjadi 1,49
mg/biji/hari. Peningkatan suhu yang lebih tinggi dari 21/16oC menjadi 30/25oC
akan mempercepat waktu pengisian biji dari 36 hari menjadi 22 hari dan laju
pertumbuhan biji meningkat dari 1,49 mg/biji/hari menjadi 1,51 mg/biji/hari (FAO
2001).
Suhu tinggi juga berpotensi mengganggu sistem perakaran tanaman
gandum karena menimbulkan efek transpiration cooling. Efek ini kemudian
berpengaruh terhadap stres panas sehingga mempengaruhi evaporasi dan
suplai air dari tanah sampai ke daun tanaman gandum. Panas yang tinggi
memicu mekanisme pergerakan air dari akar tanaman terganggu.
Remobilisasi asimilat pada tanaman gandum terjadi setelah antesis dimana
asimilat yang bersumber dari akar menjadi salah satu sumber energi utama
untuk mendukung perkembangan tanaman (Hay and Walker 1989). Terdapat
kecenderungan penurunan kerapatan panjang akar dan bobot biomas akar
akibat meningkatnya suhu karena akar bagian bawah cenderung mengalami
percepatan senescence dan meningkatnya respirasi tanaman (Ferris et al.1989).
Cekaman suhu tinggi tidak hanya mengganggu proses fisiologi tanaman
tetapi juga komposisi fisiko kimia biji gandum. Matsuki et al. (2003) menyatakan
bahwa cekaman suhu tinggi mempengaruhi kandungan pati, struktur
amilopektin, ukuran dan jumlah granula pati serta sifat gelatinisasi pati gandum.
Peningkatan suhu pada tahap pengisian biji gandum akan meningkatkan kadar
amilosa dan suhu gelatinisasi. Peningkatan suhu 15oC akan meningkatkan kadar
amilosa 1% dan suhu gelatinisasi 10,2%. Peningkatan suhu ini juga
menurunkan bobot biji 28-49%.
METODE PEMBERIAN AIR BAGI TANAMAN GANDUM
Irigasi/pemberian air tanaman dilakukan untuk menjaga kondisi lengas tanah.
Kisaran nilai kadar lengas tanah antara kapasitas lapang dan titik layu permanen
merupakan air tersedia yang dapat dimanfaatkan oleh tanaman (AW), dijadikan
indikator dalam menentukan jumlah dan waktu pemberian air. Kapasitas lapang
adalah kadar lengas tanah yang tertahan dalam tanah setelah mengalami proses
penjenuhan akibat hujan atau irigasi, yang berlangsung antara 2-3 hari setelah
hujan. Kondisi ini terjadi pada tekanan isap tanah mencapai -0,33 bar. Titik layu
permanen adalah jumlah air minimum di mana tanaman sudah mulai layu dan
tidak dapat tumbuh lagi walaupun diberi tambahan air (Een.wikipedia 2007).
Kondisi ini terjadi pada tekanan isap tanah mencapai -15 bar. Dalam prakteknya,
volume tiap satuan luas permukaan lengas tanah antara kapasitas lapang dan
Ÿc kadang-kadang disebut lengas tanah yang tersedia/siap dimanfaatkan oleh
tanaman (RAW).
Linsley dan Fransini (1986) membagi metode pemberian air bagi tanaman
ke dalam lima metode yaitu:
1. Model irigasi alur (furrow irrigation)
2. Model irigasi bawah permukaan (sub surface irrigation)
3. Model irigasi curah (sprinkler irrigation)
4. Model irigasi tetes (drip irrigation)
5. Model irigasi genangan (basin irrigation)
Di antara model ini , pemberian air dengan metode alur, irigasi tetes,
dan irigasi alur paling banyak diterapkan dalam budi daya gandum. Dengan
metode alur, air diberikan melalui alur-alur di sepanjang baris tanaman. Dengan
pemakaian alur untuk mendistribusikan air, kebutuhan pembasahan hanya
sebagian dari permukaan sehingga mengurangi kehilangan air akibat
penguapan, mengurangi pelumpuran tanah berat, dan memungkinkan untuk
mengolah tanah lebih cepat setelah pemberian air. Model irigasi pancaran dan
irigasi tetes membutuhkan air yang lebih sedikit dibandingkan dengan irigasi
permukaan. Irigasi tetes menggunakan selang kecil yang akan mendistribusikan
air pada setiap rumpun tanaman. Irigasi sprinkler disebut juga sebagai overhead
irrigation karena pemberian air dilakukan dari bagian atas tanaman terpancar
menyerupai curah hujan (Ditjen Pengelolaan Lahan dan Air Departemen
Pertanian 2008).
Ottman et al. (2012) menetapkan lima aspek untuk menentukan waktu
pemberian air irigasi (irrigation timing), yaitu tekstur tanah, kedalaman akar,
irigasi pada tahap germinasi biji, tahap generative awal, serta irigasi pada tahap soft
dough. Tekstur tanah berkaitan erat dengan kemampuan memegang air tanah.
Tanah bertekstur pasir membutuhkan air irigasi yang lebih sering dibandingkan
tanah bertekstur liat. Air tanah tersedia juga dipengaruhi oleh kedalaman akar
tanaman, dimana hampir 80-90% air tersedia diekstraksi dari akar tanaman
gandum. Irigasi juga diperlukan pada tahap germinasi biji untuk memberikan
iklim mikro yang cukup untuk menunjang pertumbuhan biji. tahap masak susu
dimana akumulasi pati telah mulai terbentuk di dalam biji juga membutuhkan
pemberian air irigasi. Irigasi yang tepat memungkinkan terjadinya akumulasi
bobot kering yang tinggi untuk menghasilkan bulir gandum yang besar dan
hasil panen tinggi (Hanson et al. 2004).
Kharrou et al. (2011) menyatakan bahwa dengan menggunakan sistem
irigasi alur, total air tersedia dalam zona perakaran akan meningkat seiring
dengan perkembangan tanaman dan akan mencapai maksimum pada
kedalaman akar 80 cm. El-Rahman (2009) dalam pengujian menggunakan sistem
irigasi tetes menunjukkan pemakaian irigasi model tetes menghasilkan efisiensi
pemakaian air yang tinggi, mencapai 1,5 kg per meter kubik air. pemakaian
sistem irigasi tetes dapat meningkatkan hasil biji gandum sebesar 28%
dibandingkan dengan sistem irigasi genangan.
Wilayah yang hanya bergantung pada curah hujan untuk pertanaman
gandum dapat menerapkan teknologi konservasi air, diantaranya menggunakan
mulsa. Zhao (1996) menyatakan bahwa konservasi air tanah sangat berperan
dalam menunjang produksi tanaman, khususnya di wilayah semi arid. Mulsa
serasah mengurangi evaporasi air tanah, meningkatkan kandungan bahan
organik dalam tanah, dan memperbaiki kualitas tanah (Langdale et al. 1992). Li
dan Lan (1995) melaporkan bahwa pemakaian mulsa plastik selain dapat
menjaga suhu tanah juga meningkatkan hasil biji gandum. pemakaian mulsa
berbahan plastik film transparan dengan ketebalan 0,007-0,008 mm mampu
menekan laju evaporasi tanah sampai 30 hari setelah tanam. pemakaian lebih
dari 30 hari tidak berpengaruh nyata terhadap produksi biji gandum (Li et al.
1999).
MODEL SIMULASI TANAMAN GANDUM
Model simulasi tanaman gandum merupakan salah satu cara untuk memahami
proses fisiologi tanaman dan memberikan rekomendasi output yang dihasilkan
berdasarkan input yang diberikan. pemakaian model simulasi tanaman
gandum memungkinkan dilakukan ektrapolasi data, memahami interaksi faktor
genetik dan lingkungan (GxE), menduga pengaruh interaksi galur terhadap
hasil dan menduga hasil ke depan pada berbagai skenario faktor lingkungan
(Ludwig and Asseng 2010; Asseng et al. 2011). Model simulasi juga
mempertimbangkan aspek ketidakpastian (uncertainties), misalnya terjadinya
hujan, suhu lingkungan, dan faktor lingkungan lainnya.
Terdapat berbagai macam model untuk menduga hasil gandum,
diantaranya CERES, APSIM wheat, AFRCWHEAT2, YIELD PROPHET, dan SIRIUS.
Jamieson et al. (1998) membandingkan akurasi model dalam menduga hasil
gandum pada berbagai tingkat cekaman air. Hasil pengujian menunjukkan
model AFRCWHEAT2 memberikan hasil yang terbaik dengan galat antara data
observasi dengan prediksi < 0,64 t/ha. Hochman et al. (2009) menggunakan
model APSIM wheat untuk menduga hasil gandum di Australia. Model ini mampu
menduga aspek fenologi tanaman, diantaranya indeks luas daun, penetrasi
akar, serapan N, dan transpirasi tanaman. Galat antara data pengamatan dengan
hasil prediksi < 0,40 t/ha.
Model simulasi tanaman gandum untuk spesifik wilayah negara kita juga
telah dikembangkan di Melbourne Australia dan divalidasi di Kuningan Jawa
Barat untuk keperluan pemetaan gandum (Handoko1994). Model ini dinamakan
model simulasi shierary wheat dan dirancang untuk mempelajari proses
interaksi antara perkembangan dan pertumbuhan tanaman dengan unsurunsur cuaca serta beberapa sifat fisik dan kimia tanah. Model ini telah divalidasi
pada skala kebun percobaan (pertumbuhan, perkembangan, neraca air, neraca
nitrogen dan produktivitas) di Kuningan pada ketinggian lokasi 600 m dpl dan
di Sulawesi Utara. Model simulasi shierary wheat mempunyai empat submodel,
yaitu (1) perkembangan tanaman/fenologi, (2) pertumbuhan tanaman, (3)
neraca air, dan (4) neraca nitrogen. Proses yang terjadi dari masing-masing
submodel saling berinteraksi menentukan pertumbuhan dan hasil
(produktivitas) gandum.
Aplikasi model simulasi tanaman gandum tidak hanya terbatas pada
pendugaan hasil berdasarkan input agronomi yang dimasukkan tetapi juga
untuk memahami keterkaitan antargen (alel gen) dengan fenotifik tanaman (G
dan F). Asseng dan Van Herwaarden (2003) mensimulasi tingkat asimilasi
menjelang tahap pengisian biji terhadap hasil biji sejumlah galur
gandum.Walaupun mampu menduga hasil dengan baik namun simulasi
mekanisme gen mengatur proses asimilasi belum dapat diduga dengan tepat.
Model ini juga belum mampu mensimulasi pengaruh perubahan alel
mempengaruhi waktu pembungaan dan hasil gandum. Pada gandum musim
dingin, data sebagian alel Ppdisoline telah berhasil di dapatkan (Worland 1996;
Foulkes et al. 2004; González et al. 2005). Saat ini, model simulasi interaksi genetiklingkungan masih menjadi tantangan terbesar bagi pakar pemodelan tanaman.
Tanaman gandum (Triticum aestivum L.) bukan merupakan tanaman daerah
tropika, namun dikenal sebagai tanaman yang berasal dari Asia Barat beriklim
mediteran kering (Klages 1958). Daerah budi daya gandum, Curtis (1988)
membagi negara-negara tropika menjadi dua tipe lingkungan utama, yaitu (1)
wilayah dengan suhu udara tinggi, kelembaban udara rendah, periode tumbuh
pendek dan sedikit ditemukan permasalahan penyakit tanaman, dan (2) wilayah
dengan suhu dan kelembaban udara tinggi, perode tumbuh pendek, dan
penyakit tanaman lebih dominan.
negara kita digolongkan ke dalam tipe lingkungan kedua, walaupun
sebenarnya kedua tipe lingkungan utama ini terdapat di negara kita .
Kawasan Timur negara kita lebih mewakili tipe lingkungan yang pertama,
sedangkan Kawasan Barat negara kita mewakili tipe lingkungan kedua (Zaini et
al. 1991). Sesuai dengan letak geografis negara kita di daerah tropis, maka jenis
gandum yang mungkin dibudi dayakan adalah gandum musim dingin (winter
wheat). Jenis gandum ini dapat tumbuh pada kisaran suhu udara antara
4-31oC, dengan suhu udara optimum rata-rata 20o
C (Fisher 1983).
Tulisan ini membahas kesesuaian lingkungan, pendekatan fisiologis dan
agronomis dalam pengembangan gandum di daerah dengan cekaman suhu
tinggi.
KESESUAIAN LINGKUNGAN TANAMAN GANDUM
Curah Hujan
Tanaman gandum membutuhkan air dan kelembaban udara yang relatif lebih
rendah dibandingkan dengan tanaman serealia lainnya. Curah hujan terbaik
untuk tanaman gandum berkisar antara 640-890 mm/tahun. Diperlukan adanya
bulan kering (<100 mm/bulan) pada periode akhir sebelum panen. Curah hujan
yang tinggi pada periode satu bulan sebelum panen akan meningkatkan
penularan penyakit scab oleh Fusarium spp. di samping menyukarkan
perontokan gabah dan menurunnya kualitas tepung yang dihasilkan.
Kassam (1976) dan FAO (1978) mengemukakan kesesuaian lingkungan/
iklim untuk tanaman gandum adalah laju fotosintesa 20-30 mg CO2
/dm2
/jam,
suhu udara rata-rata 15-23o
C, radiasi surya 0,2-0,6 kal/cm2
/menit, dan curah
hujan 60-155 mm/bulan.
Suhu Udara
Suhu udara (maksimum dan minimum) merupakan faktor utama dalam
menentukan kesesuaian wilayah untuk tanaman gandum. Tanaman gandum
memberikan produksi optimal pada kisaran suhu udara 10-20oC (Fisher 1985)
dan kisaran ini hanya terdapat pada daerah-daerah di sekitar lintang 20o
atau lebih tinggi. Di sekitar garis khatulistiwa, kisaran ini sangat dipengaruhi
oleh tinggi tempat dari muka laut. Di negara kita , kisaran suhu ini ditemukan
di daerah dengan ketinggian lebih dari 500 m di atas muka laut.
Aggarwal (1991) menggunakan model simulasi dalam memprediksi
produktivitas gandum yang dikaitkan dengan suhu udara. Untuk setiap derajat
celcius kenaikan suhu udara rata-rata, hasil gandum akan turun 504 kg/ha. Di
daerah dengan suhu udara rata-rata 22,5-27,5oC hasil yang dapat dicapai hanya
1,9-4,4 t/ha, pada suhu 17,5-22,5oC memberi hasil 4,5-6,9 t/ha, dan pada suhu
12,5-22,5oC diperoleh hasil 7,0-9,4 t/ha. Rendahnya potensi hasil pada daerah
berelevasi rendah erat kaitannya dengan respon tanaman terhadap suhu udara.
Suhu yang tinggi memperpendek tahap vegetatif tanaman, sehingga potensi
genetik tanaman tidak tercapai (Kramer 1980).
Menurut Reynolds et al. (2001), penanaman gandum di wilayah tropika
mempunyai kelemahan, terutama cekaman suhu tinggi yang mempengaruhi
pertumbuhan tanaman. Cekaman panas terus-menerus didefinisikan sebagai
suhu rata-rata harian lebih dari 17,5°C pada bulan paling dingin dalam satu
musim (Fokar et al. 1998). Lebih dari 50 negara (yang mengimpor lebih dari 30-
40 juta ton gandum per tahun) mengalami cekaman ini sepanjang siklus
tanaman gandum. Hal ini memicu sebagian besar negara-negara
berkembang di daerah tropika memilih cekaman suhu tinggi sebagai salah satu
prioritas penelitian utama mereka (Reynolds et al. 2001).
Kerja sama penelitian International Heat Stress Genotype Experiment (IHSGE)
antara CIMMYT dan NARS pada aspek fisiologis cekaman suhu tinggi tanaman
gandum dimulai pada tahun 1990, melibatkan peneliti gandum di Bangladesh,
Brazil, Mesir, India, Nigeria, Sudan, dan Thailand (Reynolds et al. 1998). Tujuan
utama IHSGE adalah untuk mengamati keragaman genetik dalam kaitannya
dengan suhu tinggi, dan meningkatkan pemahaman tentang dasar-dasar
fisiologis dan genetika dari cekaman suhu tinggi.
Hasil utama penelitian IHSGE dalam periode 1990-94 menunjukkan: (1)
interaksi antara lingkungan dan genotipe dipengaruhi oleh kelembaban udara
relatif (RH), baik pada negara-negara yang mempunyai RH rendah (misalnya
Sudan, Meksiko, dan India) maupun RH tinggi (seperti Bangladesh dan Brasil),
dan (2) terdapat hubungan yang konsisten antara hasil gandum dan beberapa
komponen morfologi tanaman (Tabel 1).
KARAKTER FISIOLOGIS TERKAIT TOLERANSI PANAS
Keanekaragaman genetik untuk toleransi panas dalam budi daya gandum telah
banyak diketahui (Midmore et al. 1984, Rawson 1986, Wardlaw et al. 1989; AlKhatib and Paulsen 1990, Reynolds et al. 1994). Cossani dan Reynolds (2012)
mengemukakan jika air dan nutrisi tidak menjadi faktor pembatas, hasil gandum
menjadi fungsi dari tiga karakter utama secara genetis, yaitu: (1) intersepsi cahaya
(LI); (2) efisiensi pemakaian radiasi (RUE); dan (3) partisi total asimilasi.
Asimilasi fotosintesa menjadi faktor pembatas di bawah tekanan cekaman
suhu tinggi di wilayah tropika dibandingkan dengan lingkungan beriklim sedang,
terutama karena cekaman suhu tinggi meningkat selama proses pengisian biji.
Pada saat yang sama, kebutuhan hasil asimilat meningkat cepat. Hal ini
diindikasikan oleh pengamatan bahwa di bawah cekaman suhu tinggi, jumlah
biomassa di atas tanah berkorelasi sangat nyata dengan hasil biji dibandingkan
dengan indeks panen (Tabel 1). Situasi ini biasanya terbalik pada wilayah beriklim
sedang. Oleh karena itu, karakter yang mempengaruhi efisiensi pemanfaatan
radiasi matahari, seperti persentase penutupan tanah awal, tetap hijau/stay
green, dan laju fotosintesis, menjadi penting di bawah tekanan suhu tinggi.
Bukti fisiologis menunjukkan bahwa menurunnya jumlah klorofil selama proses
pengisian biji berkaitan erat dengan penurunan hasil gandum di lapangan
(Reynolds et al. 1994).
Studi dalam lingkungan terkendali (growth chamber) mengungkapkan
adanya variabilitas genetik pada tingkat fotosintesis antara kultivar gandum bila
terkena cekaman suhu tinggi (Wardlaw et al. 1980, Blum 1986). Perbedaan
fotosintesis di bawah cekaman suhu tinggi terkait dengan menurunnya jumlah
klorofil dan perubahan dalam rasio klorofil a:b karena proses penuaan daun
yang lebih awal (Al-Khatib and Paulsen 1990, Harding et al. 1990, Moffat et al.
1990). Penurunan suhu kanopi (canopy temperature depression/CTD),
ketahanan stomata pada daun bendera, dan laju fotosintesis berhubungan
dengan keragaan tanaman di lapangan di bawah cekaman suhu tinggi (Reynolds
et al. 1994). Ketahanan stomata dan CTD merupakan mekanisme melepaskan
diri (escape mechanism) dari cekaman suhu tinggi (Amani et al. 1996, Shanahan
et al. 1990).
Menggunakan konduktimeter untuk mengukur kebocoran zat terlarut dari
sel, diperkirakan kerusakan akibat cekaman suhu tinggi terjadi pada membran
plasma atau membran termostabilitas (MT) (Blum and Ebercon 1981). Saadalla
et al. (1990) menemukan korelasi yang tinggi antara kerusakan MT pada stadia
bibit dan stadia generatif (daun bendera) untuk genotipe yang tumbuh di bawah
kondisi lingkungan terkontrol yang diberi perlakuan cekaman suhu tinggi.
Hilangnya integritas membran mungkin menjadi penyebab kebocoran ion
sel. Fenomena ini juga bisa disebabkan oleh suhu tinggi yang mendorong
penghambatan aktivitas enzim yang terikat pada membran yang berfungsi
memelihara keluar masuknya ion-ion di dalam sel. Bukti langsung terjadinya
penghambatan proses biokimia pada mekanisme toleran suhu tinggi pada
tanaman gandum adalah tidak aktifnya enzim pelarut pati synthase pada suhu
tinggi (Keeling et al. 1994). Konversi sukrosa menjadi pati dihambat pada kondisi
cekaman suhu tinggi, yang menjelaskan peningkatan kadar karbohidrat dalam
jaringan vegetatif terjadi pada proses pengisian biji (Bhullar and Jenner 1986,
Gutierrez-Rodriguez et al. 2000).
Mekanisme fisiologis yang berbeda dapat berperan dalam toleransi suhu tinggi
kultivar gandum di lapangan, seperti ditunjukkan oleh tingkat fotosintesis yang
lebih tinggi, daun tetap hijau, membran thermostability, atau menghindari panas
seperti yang ditunjukkan oleh penurunan suhu kanopi. Program pemuliaan
dapat mengukur sifat-sifat ini untuk membantu pelaksanaan seleksi galurgalur toleran suhu tinggi.
Data lapangan menunjukkan hubungan yang nyata antara CTD dengan
hasil biji, baik pada lingkungan suhu tinggi maupun beriklim sedang. CTD
menunjukkan korelasi genetik yang tinggi dengan hasil biji. Reynolds et al. (1998)
melaporkan sifat ini diwariskan sehingga dapat digunakan dalam seleksi
generasi awal. Nilai CTD dapat diukur hampir seketika (real time) pada sejumlah
tanaman dalam petak pemuliaan yang kecil. CTD dipengaruhi oleh banyak faktor
fisiologis, yang membuatnya menjadi sifat integratif yang kuat, tetapi
pemakaian nya mungkin dibatasi oleh kepekaan terhadap faktor lingkungan.
Pengukuran CTD pada 60 galur harapan tanaman gandum di Meksiko
menunjukkan korelasi yang tinggi dengan hasil biji (Tabel 2).
Penurunan suhu kanopi tanaman sangat sesuai untuk seleksi galur-galur
unggul secara fisiologis pada lingkungan suhu tinggi dengan kelembaban relatif
yang rendah, evaporasi tinggi yang memicu pendinginan daun hingga
10°C di bawah suhu kamar. Perbedaan di antara genotipe untuk dideteksi relatif
mudah dengan menggunakan thermometer inframerah. Namun, perbedaan
ini tidak dapat dideteksi dalam lingkungan kelembaban relatif tinggi, karena
efek pend






.jpg)



